Egy új sejttípus borzolja manapság az immunológusok idegeit; a jövevény neve a keresztelôben az IKDC nevet kapta (interferon-producing killer dendritic cell, azaz interferon-termelő, ölő dendritikus sejt).
De miért is érdekes ez? Az új sejttípus az immunrendszer egyfajta kacsacsőrű emlőse, ami magán hordozza több, jól ismert sejt tulajdonságát. Mely sejtekre hasonlítanak tehát az IKDC-k? Mint, ahogy az a Nature Medicine legújabb számában megjelent két független kutatócsoport eredményeiből kiderül, az új kentaur sejttípus egyszerre dendritikus sejt es termeszétes ölősejt (natural killer, azaz NK sejt) is egyben. [1,2]
Ezen sejttípusok már régóta ismertek az immunológusok számára. A dendritikus sejtek (DC, dendritic cell) a szervezet őrszemei. A szövetekben lazítanak, ahol a kórokozókat (pl. baktériumokat) észlelve, azokat felveszik és a nyirokcsomókba szállítják. Itt a kórokozókból származó fehérje darabokat T sejteknek mutatják be, s a fehérje fragmenseket specifikusan felimerő T sejtek pedig az interakció következtében aktiválodnak ( ezt nevezzük klonális szelekciónak), azaz beindul az immunválász. A buli végén – többek közt – a B sejtek által termelt ellenanyagok segítségevel, a kórokozó eltávolítódik a szervezetből. Ez az ún. adaptív immunitás; nagy specificitású ellenanyagok termelése és immun-memória kialakulása jellemzi (ezen a jelenségen alapulnak a vakcinációk is). Hátránya, hogy viszonylag hosszú időt, mintegy két hetet vesz igénybe. Tekintve a bakteriumok gyors osztodási sebességet (akár 20 perc) ez önmagában nem lenne elegendő a boldogsághoz. Szerencsére segítségünkre van egy ennel sokkal ôsibb rendszer is; a természetes immunitás. Ennek a részei a falósejtek, (makrofágok) természetes ölősejtek (natural killer, vagyis NK sejtek) illetve egy, a vérben jelenlévő nagy hatékonyságú enzimrendszer, a komplement rendszer. A természetes immunitás evolúciósan sokkal korábban jelent meg, már a rovarokban is megtalálhatók egyes sejttípusok, illetve molekulák ősei. Ezzel szemben az adaptív immunitás először a halakban jelent meg, bár nemrég egy hasonló rendszert írtak le a gyümölcslégyben (Drosophila melanogaster) is. A természetes immunitás setjei és molekulái azonnal reagálnak a kórokozókra, mintegy első vonalbeli védelmet biztosítva.
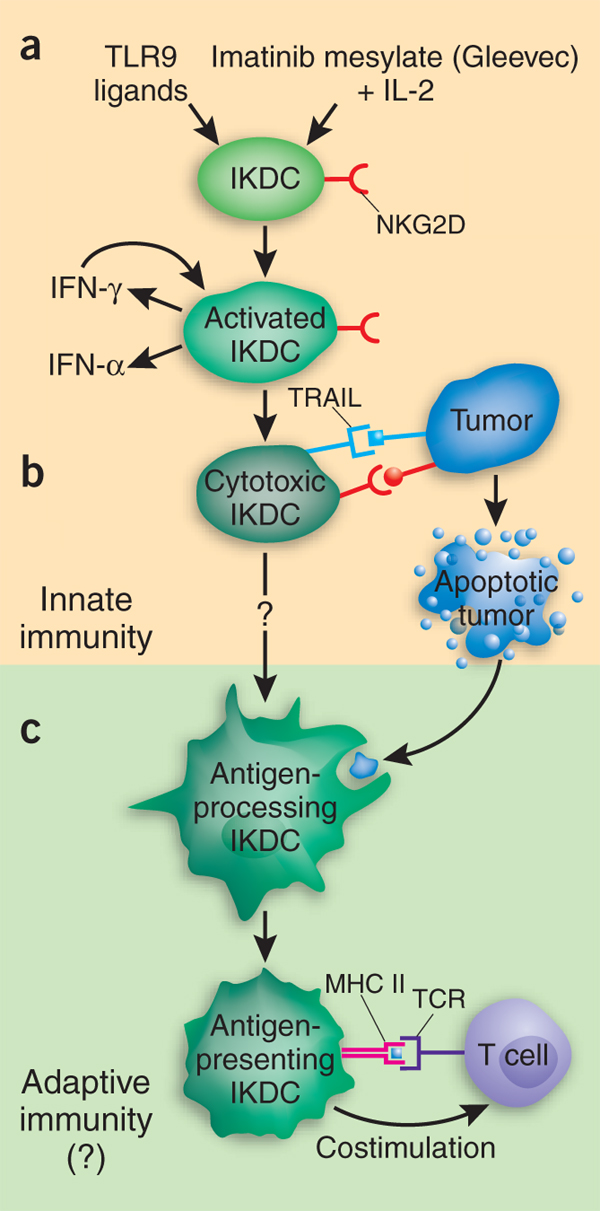
Térjünk vissza az IKDC-hez. Ezek a sejtek kapcsot alkotnak az immunrendszer két ága között: képesek nagy mennyiségben interferon-alfá-t termelni (plamacytoid DC tulajdonság), sejteket megölni a TRAIL nevu halál-receptoron keresztul, (NK sejt tulajdonság) illetve antigéneket prezentálni T sejteknek (myeloid DC tulajdonság). Az NKDC sejtek igen homogén populációt alkotnak, több sejtfelszíni molekula expressziója szerint azonosíthatók. Sok kutató számára bizonyosan egyes DC-kkel illetve NK sejtekkel végzett kisérletek során kapott „furcsa” eredményekre is magyarázatot adhat az IKDC-kkel való szennyezôdés.
Felmerül a kérdés; mi lehet evoluciósan a jelentősége egy olyan sejtnek, ami egyesíti más, hasonló feladatokra szakosodott sejtek funkcioit? Az egyik elmélet szerint az új sejt fő szerepe a tumorok elleni védekezés (tumor surveillance). Könnyű belátni, ha egy sejt képes felismerni és megölni a tumorsejteket, ugyanakkor azonnal képes is felvenni és feldolgozni a tumor eredetű fehérjéket, majd azokkal indukákni a T sejteket (ezáltal hatékony adaptív immunválaszt váltva ki), akkor az a sejt igazán legény a gáton.
Számos bizonyíték van rá, hogy a természetes és az adaptív immunrendszer nem két egymástól elválasztható, független rendszer, hanem egymással több ponton állandó kapcsolatban álló, sok kölcsönös visszacsatolással rendelkezô dinamikus hálozat. Az új sejttípus létezése ennek egy újabb ékes bizonyítéka. Az IKDC-ket a tumor immunterápiában való ígéretes szerepük miatt várhatóan több csoport is górcső alá fogja venni, ezert minden bizonnyal még hallunk róluk a jövőben is.
[1] Interferon-producing killer dendritic cells provide a link between innate and adaptive immunity
Camie W Chan, Emily Crafton, Hong-Ni Fan, James Flook, Kiyoshi Yoshimura, Mario Skarica, Dirk Brockstedt, Thomas W Dubensky, Monique F Stins, Lewis L Lanier, Drew M Pardoll & Franck Housseau
Nature Medicine, 29 January 2006, pp207 – 213
[2] A novel dendritic cell subset involved in tumor immunosurveillance
Julien Taieb, Nathalie Chaput, Cédric Ménard, Lionel Apetoh, Evelyn Ullrich, Mathieu Bonmort, Marie Péquignot, Noelia Casares, Magali Terme, Caroline Flament, Paule Opolon, Yann Lecluse, Didier Métivier, Elena Tomasello, Eric Vivier, François Ghiringhelli, François Martin, David Klatzmann, Thierry Poynard, Thomas Tursz, Graça Raposo, Hideo Yagita, Bernard Ryffel, Guido Kroemer & Laurence Zitvogel
Nature Medicine, 29 January 2006, pp214 – 219

 Már a 19. században számos anatómus felfigyelt arra, hogy a csirkék embrionális fejlődése során átmeneti duzzanatok (papillák) jelennek meg a fejlődő embrió állkapcsán, amelyek sokban emlékeztetnek hüllő fogakra. Sőt, mint arról
Már a 19. században számos anatómus felfigyelt arra, hogy a csirkék embrionális fejlődése során átmeneti duzzanatok (papillák) jelennek meg a fejlődő embrió állkapcsán, amelyek sokban emlékeztetnek hüllő fogakra. Sőt, mint arról