Na akkor jöjjön a testalkathoz kapcsolódó Nature Review Genetics cikkek közül az utolsó (pontosabban az újságban ez állt legelöl, de nekem jobban feküdt, hogy a végére hagyjam ;-)) [1].
A test felosztásában és egyes végtagok, specifikus csigolyák, stb.
helyzetének meghatározásában kulcs szerepet játszanak a
Hox-gének, amelyek, mint arról már szó volt,
nemcsak az expressziójuk során, de a kromoszómákon is egymás után
helyezkednek el. Ennek minden valószínűség szerint azon egyszerű
evolúciós oka van, hogy ezek a gének mind közös tőről szakadnak, azaz
egy ősi ProtoHox gén duplikációiból származó négytagú Proto-Hox
csoport (cluster) egyes tagjaira vezethetők vissza. Ami érdekes, ennek
a csoportnak a duplikációja során létre jött még egészen régen egy
úgynevezett ParaHox cluster is. Ez az a géncsoport kevesebb génből
áll, és ezek egy másik csíralemezben expresszálódnak mint a Hox-gének,
sőt a gerinctelenekben nem is együtt találhatók, de az egyik legősibb
gerincesnek tartott lándzsahal kromoszómáin szintén egymás mellett
vannak. Vagyis valószínűsíthető, hogy a ma ismert Hox és Hox-rokon
gének a kezdeti tandem duplikációk után fizikailag kettéváltak, azaz
külön kromoszómákra kerültek.
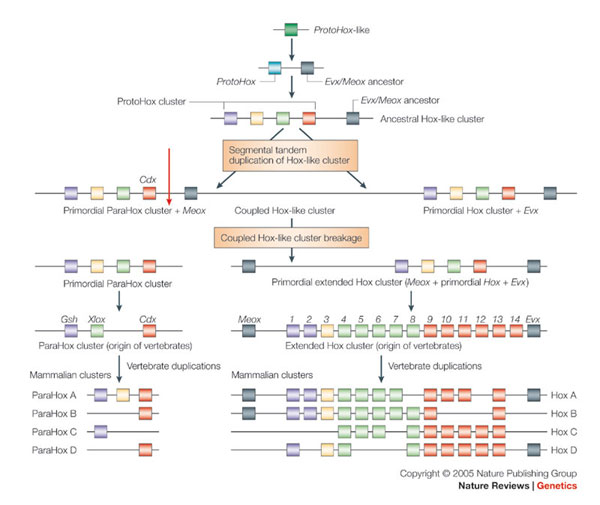
Érdekesség, bár ezen az ábrán nincs feltüntetve, hogy a Hox- és ParaHox
csoportok mellett létezik egy harmadik rokon csoportosulás, az NK gének
csoportja is. Ezek már szintén kissé szétszórva lelhetők fel a
különböző genomokban, de ha végigböngésszük a már rendelkezésünkre álló
szekvenciákat, akkor a csoportot alkotó egyes gének különböző
kombinációkban és különböző fajokban, de előfordulnak együtt, azaz
egykori kolinearitásuk szintén valószínűsíthető. (Természetesen ez
felvet egy érdekes, és mindeddig még megválaszolatlan kérdést: miért
maradtak együtt a Hox-gének, ha a rokon csoportok tagjai ennyire
szétszóródtak?) Hogy még érdekesebb legyen a dolog, míg a Hox gének az
ektodermában (a bőrt, idegrendszert és arckoponyát kialakító
csíralemezben), a ParaHox gének az endodermában (az emésztőrendszer nagyobb részét és
légzőrendszert, valamint endokrin mirigyeket létrehozó csíralemezben)
fejeződnek ki, addig az NK gének, a harmadik csíralemezben, a
mezodermában lelhetők. Ebből aztán adódik az igen csábító (de még nem
bizonyított) elmélet, hogy valamilyen köze lehetett az ősi ProtoHox gén
duplikációinak a csíralemezek kialakulásához. Jól hangzik, de van benne
egy kis gubanc: mindhárom kluszter megtalálható már a csalánozókban is,
márpedig ezek az állatok a klasszikus felfogás szerint
diploblasztikusak, azaz csak két csíralemezük van (az ekto- és az
endoderma).
A paradox helyzet azonban nem feloldhatatlan. A jelenleg elterjedt
felfogás szerint, a két csíralemezből álló és szimmetriájú csalánozók a
kétoldali szimmetria és három
csíralemezű állatok őseinek tekinthetők, és utóbbiak két
kulcstulajdonsága egyszerre jelent meg az evolúció során. Azonban,
mint a
sorozat első részében említettem,
a legújabb kutatások tükrében ez nem evidens. Ugyanis egyre több
csalánozóról derül ki, hogy bár kívülről körkörös szimmetria fedezhető
fel bennük, de génexpressziós szinten kétoldali szimetriájúak, vagyis
van olyan gén ami az állat egyik oldalán kifejezésre kerül, de a másik
oldalon nem. Sőt vannak olyan csalánozó fajok, ahol egyértelmű, hogy
vannak olyan izomszerű sejtek amelyek csakis a harmadik csíralemezből
származhatnak. Summa sumarum még az is lehet, hogy a körkörös szimmetria (már ha valóban létezik) nem
ősi, hanem csak később létrejövő tulajdonság, és maguk a csalánozók is
szélsőségesen módosult kétoldali szimmetriájú állatok. Márpedig ilyen
szélsőséges módosulás együtt járhatott a harmadik csíralemez
módosulásával és elvesztésével, vagyis a Hox, ParaHox és NK clusterek
jelenléte ebben a csoportban nem mond ellent annak, hogy eredetileg
közük volt a három különböző csíralemez létrejöttéhez.
 És akkor végül elérkeztünk a legfogasabb kérdéshez: mi volt a szerepe
És akkor végül elérkeztünk a legfogasabb kérdéshez: mi volt a szerepe
az ősi ProtoHox génnek? Mint sok helyen máshol itt is egy kicsit
találgatásokra vagyunk utalva, de azért van egy reménysugár, ami elég
meglepő helyről érkezi. Vagy nem is, ha jobban belegondolunk: a
legprimitívebb többsejtű állatnak tartott Placozoákról van szó, amelyek
nem többek egy többrétegű mikroszkopikus, lapos és mozgó nyálkafoltnál (lásd jobboldalt).
Ezeknek az állatoknak sem bélürege, sem csíralemezei nincsenek, és a
többi állatra jellemző bazális lamina, valamint a sejtek közötti teret
kitöltő ún. extracelluláris mátrix sem fedezhető fel bennük. Mindössze
néhány sejttípus lelhetünk, amelyek aszimmetrikusan helyezkednek el: az
alsó rétegben lelhetők a táplálék felvételéért felelős epiteliális
sejtek. (Minden primitívségük ellenére, még azt sem tudjuk, hogy
szaporodnak-e ezek az állatok szexuálisan (laborban csak valami
osztódás szerűságet tudtak megfigyelni), és ha igen, milyen környezeti
hatások erdményeként hoznak létre ivarsejteket.)
Nos, ebben a bizarr kis lényben egyetlen Hox-gént találtak (amit valamiért Trox-2
névre kereszteltek), és ez a gén az állat szélein expresszálódik [2]. Azaz
mint az sejthető volt eredetileg nem a hosszanti tengely kialakításában
játszottak a Hox-gének szerepet (mert ilyen tengely nem is volt), hanem
más dolguk volt. (Majd később kooptálódtak, ugye, a mai feladatuk
betöltésére ;-)). Már csak arra kellene fényt deríteni, hogy pontosan
mi is. Egyelőre elég kevesen bogarászták ezeket a furcsa kis dögöket,
de a nemrég elindult genom-projecteknek köszönhetően az érdeklődés
hamarosan valszleg nőni fog, így ilyen szaftos részletekre is fény
derülhet.
[1] Garcia-Fernandez J (2005) The genesis and evolution of homeobox gene clusters. Nat Rev Genet. 6(12): 881-892.
[2] Schierwater B (2005) My favorite animal, Trichoplax adherens. Bioessays 27:1294-1302.
Előzmények: Test, alkat – 1. | Test, alkat – 2