
A már említett tavaly decemberi Nature Reviews Genetics
melléklet egy másik cikke a testfelépítés létrejöttében kulcsszerepet
játszó homeobox (Hox) génekkel foglalkozik, pontosabban azzal, hogy
miként is variálódhat ezek funkciója.[1]
A Hox gének a test hosszanti felosztásáért felelnek, expressziójuk
(illetve különböző Hox gének expressziójának kombinációja)
meghatározza, hogy adott pozícióban milyen struktúrák (pl. végtagok)
fejlődnek. Két igen érdekes tulajdonságuk van: egyrész a többsejtű
állatokban szinte univerzálisak és ugyanazt a funkciót látják el (1.
Ábra), másrészt a kromoszómákon egymás mellett találhatóak és
jellegezetes időbeli és térbeli kollinearitást mutat a genomban való
elhelyezkedésük: vagyis az a gén amelyik a „sorban” előrébb található
általában hamarabb és a fejhez közelebbi területen jut kifejeződésre.

1. Ábra: A homebox gének mind a testtengely
mentén való expressziója, mind a kromoszómákon való helyzete igen
hasonló a legkülönbözőbb fajok között.[1]
Annak ellenére, hogy rengetegen foglalkoznak ezekkel a génekkel,
hosszú évek óta, a szabályozásukról illetve a kollinearitás okáról még
mindig csak halvány sejtéseink vannak. Tudjuk, hogy az egyes Hox gének
képesek egymás expresszióját szabályozni, és nemrég arra is fény
derült, hogy rövid, nem kódoló, de egyes gének végső szakaszával
komplementer mRNS molekulák (miRNA) is szabályozhatják a
kifejeződésüket (ezek a génről átíródott mRNS-hez kötődve serkentik
annak lebontását) (2. Ábra). A kollinearitás oka egyesek szerint a
kromoszómák, pontosabban az ún. kromatin szerkezetében kereshető. A DNS
molekula ugyanis jellegzetes módon ösze van csomagolva, és általában
csak az éppen átíródó gének körül lazul fel ez a csomagolás (az ilyen
laza csomagolódású DNS alkotja az ún. eukromatint). Mivel a Hox gének
fizikailag egymás után találhatók a kromoszómán, elképzelhető, hogy
amikor ezen gének átírásra kerülnek a csomagolás legelől nyílik először
fel, és az eukromatinná való alakulás csak fokozatosan terjed tova, ami
időbeli eltolódást okoz a hátrébb lelhető gének kifejeződésében.
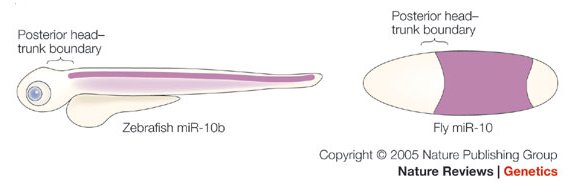
2. Ábra: A homebox gének szabályozása is
evolúciósan konzervált: mind a zebrahal, mind a muslica fejlődő
egyedeiben igen hasonló expressziója miRNA molekulák szabályozzák egyes
Hox gének átírásának helyét.[1]
A Hox gének expressziójának bütykölése az evolúció kedvelt időtöltése, mint azt láttuk a kígyók esetében,
de számos egyéb példa is akad: az egyik legelegánsabb a rákfélék
csoportjához kapcsolható, ahol kimutatták, hogy az Ubx-Antp homeobox
gének expressziója befolyással van arra, hogy egyes rendekben a torhoz
kapcsolódó végtagok az étkezésben használatos szervé ún. maxillapédiává
módosulnak vagy lábként működnek (3. Ábra).[2]
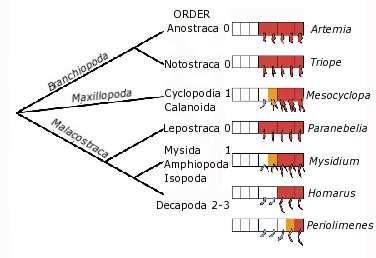
3. Ábra: A rákfélék között az Ubx-Antp gének
expressziója pontosabban annak hiánya szabályozza a tor egyes
szelvényein lelhető nyúlványok identitását: ahol az említet gének
hiányoznak (fehér) vagy csak igen alacsony szinten vannak jelen
(narancssárga), ott a táplálkozásnál használt maxillapédiák alakulnak
ki, szemben a magas expressziójú szelvényekkel (piros), ahol lábakat
találunk.[2]
Végezetül ugyancsak az ízeltlábúak jóvoltából, arra is tudunk példát
mutatni, hogy homeobox gén elveszti a homeotikus funkcióját és más
szerepet kap. A Drosophila bicoid (bcd) génje például
gazdája hossztengelyének legfontosabb szabályozója: már az mRNS is a
pete egyik (leendő elülső) végében koncentrlódik, amely révén egy
gradiens jön létre a fehérje szinten. Ez a gradiens szabályozza aztán
később más, a testfelépítésben fontos transzkripciós faktorok
működését. Azonban, mint az már régebben kiderült ez egy kicsit különc
tulajdonsága a muslicáknak és a legtöbb többi rovarban más rendszer
felelős a fent leírt szabályozásért. A bcd eredetileg egy Hox3
gén duplikációjával jött létre (erről helyzete is árulkodik, 1. Ábra),
majd fokozatosan elvesztette eredeti funkcióját és újat vett fel
(részletesebben a Pharyngulán olvashat erről, akit érdekel).
A másik példa, szintén Drosikból a fushi-tarazu (ftz)
gén esete, amely a Hox6 génből származtatható, és amely fokozatos
doméncsereberével egy klasszikus homebox génből, egy a szegmentációt
befolyásoló génné alakult (4. Ábra) [3,4].

4. Ábra: Az ízeltlábúakon belül jól követhető amint a fushi-tarazu gén homeotikus génből, szegmentációt szabályozóvá válik.[4]
[1] Pearson JC, Lemons D, McGinnis W. (2005) Modulating Hox gene functions during animal body patterning. Nat Rev Genet. 6(12): 893-904.
[2] Averof M, Patel NH. (1997) Crustacean appendage evolution associated with changes in Hox gene expression. Nature 388: 682-6.
[3] Damen WG. (2002) fushi tarazu: a Hox gene changes its role. Bioessays 24(11): 992-5.
[4] Lohr U, Pick L.
(2005) Cofactor-interaction motifs and the cooption of a homeotic Hox
protein into the segmentation pathway of Drosophila melanogaster.Curr Biol. 15(7):643-9.
Előzmény: Test, alkat – 1.