A külvilágból beérkező információink 70%-át a szemünkön keresztül szerezzük, nem véletlenül tartjuk sokra látórendszerünket. De sajnos, ami elromolhat, az el is romlik, így számos olyan betegség ismert (az esetleges baleseti sérüléseket nem is említve), amely pont ezt a finom érzékszervünket teszi gallyra.
Ezen betegségek egyik nagy csoportja, a fény érzékelésért elsődlegesen felelős ún. fotoreceptor sejteket (emlékezzünk csak: csapok és pálcikák) érinti, és a tudomány jelen állása szerint ez (sajnos) lassan, de biztosan a beteg megvakulásához vezet. A tudomány állása azonban változni látszik, ui. úgy tűnik, hogy sejtbeültetéssel sikeresen lehet kezelni a betegség egérmodelljeit.
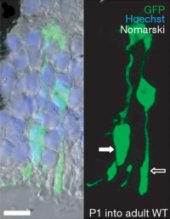
Az ezt bemutató Nature cikk, azért is tanulságos, mert nagyon jól nyomon követhető a kutatás logikája is. Először olyan újszülött (P1) kisegerek retinájából vettek kis sejteket (az egerek retinájában a pálcikasejtek fejlődésének utolsó szakasza ekkor zajlik), amelyek egy zöld fluoreszcens fehérjét expresszálnak sejtjeikben (így követni lehet, hogy mely sejtek származnak a donorból) és ezeket a transzgént nem hordozó P1 társaik szemébe ültették. A sejtek szépen integrálódtak az új gazda retinájába, a várakozásnak megfelelően főleg pálcika sejteket hozva létre, de néha akadt egy-két csapsejt is (csak emlékeztetőül: utóbbiaknak a nappali színlátásban van fontos szerepe, míg előbbieknek a szürkületi és esti félhomályban való tájékozódásban).
Mivel a P1-es stádiumban az egerek szemében sokfajta sejttípus előfordul őssejtektől kezdve, kifejlett fotoreceptorokig, azt próbálták meghatározni a következő lépésben, hogy pontosan melyik sejtek integrálódnak sikeresen. A várakozás, talán mondanom sem kell, az volt hogy multipotens őssejtek lesznek a hunyók, de meglepetésre éppen hogy a már elkötelezett, de még nem teljesen kifejlődött fotoreceptorok bizonyultak a legjobb átültetési alanyoknak.
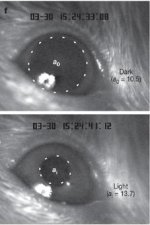
Következő lépésben, a kisegerek retina sejtjeit előbb egészséges- , majd retina degenerációban szenvedő felnőtt egerekbe ültették. Mindkét esetben a sejtek szépen integrálódtak a retinába, megfelelő szinaptikus kapcsolatokat hoztak létre a szomszédos sejtekkel, és minden jel szerint jól végezték a feladatukat. Míg beteg egerek kontroll, azaz kezeletlen szemében kis fényerősségen egyáltalán nem lehet megfigyelni a pupilla összehúzódását, addig az átültetéssel kezelt szemekben (főleg ha nagyszámú sejt integrálódott) a pupilla-reflex sikeresen kiváltható volt így is. (Azaz a sejtekrendeltetésszerűen továbbították a beérkező információt.)
Az eljárás orvosi vonatkozásait talán fölösleges is ecsetelnem, de erre még valószínűleg várni kell egy kicsit. Előbb ugyanis azt kell megfejteni, miképpen lehet sejttenyészetekben előállítani a transzplantációra alkalmas stádiumú fotoreceptorokat. Mert a kisegerekből való sejtizolálás humán megfelelőjét nem nevezém járható útnak.
MacLaren, RE, Pearson, RA, MacNeill, A, Douglas, RH, Salt, TE, Akimoto, M, Swaroop, A, Sowden, JC, Ali, RR (2006) Retinal repair by transplantation of photoreceptor precursors. Nature 444: 203-207.

 Ha a nemek mögött levő genetikai faktorokról beszélünk, elsősorban az X és Y kromoszómára gondolunk. Pedig az emlősökre olyan jellemző „XY” szexdeterminációs rendszer (egyébként vannak kivételek, azokról majd máskor) csak logikájában tekinthető viszonylag elterjedtnek az állatvilágban, mechanizmusában egyáltalán nem.
Ha a nemek mögött levő genetikai faktorokról beszélünk, elsősorban az X és Y kromoszómára gondolunk. Pedig az emlősökre olyan jellemző „XY” szexdeterminációs rendszer (egyébként vannak kivételek, azokról majd máskor) csak logikájában tekinthető viszonylag elterjedtnek az állatvilágban, mechanizmusában egyáltalán nem.

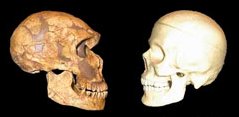 Az emberiség bölcsője Afrika, méghozzá (a jelenleg legelfogadottabb elmélet szerint) többszörösen is. A kétlábra emelkedő Homo genus először úgy 1 millió éve kolonizálta az eurázsiai kontinenst a Homo erectus formájában, majd ezt újabb hullámok követték. Először fél millió éve a Homo heidelbergensis egy csoportja kelt útra, majd úgy 75-100,000 éve a modern ember őse, a Homo sapiens.
Az emberiség bölcsője Afrika, méghozzá (a jelenleg legelfogadottabb elmélet szerint) többszörösen is. A kétlábra emelkedő Homo genus először úgy 1 millió éve kolonizálta az eurázsiai kontinenst a Homo erectus formájában, majd ezt újabb hullámok követték. Először fél millió éve a Homo heidelbergensis egy csoportja kelt útra, majd úgy 75-100,000 éve a modern ember őse, a Homo sapiens.
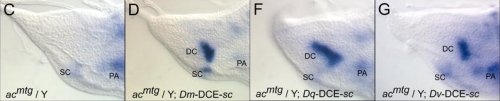
 A tengeri sünök kétségtelen, hogy messze nem a legfontosabb és legnépszerűbb állatok (tengerparton ejtőzők szimpátiaindexén valahol egy homokba fúródott üvegcserép és egy különösen éles kődarab között helyezkednek el), de megvan a maguk helye mind a tengerek ökoszisztémáiban, mind a laboratóriumok asztalain. Előbbiekben algával táplákozó primer konzumensek, amelyeket a tápláléklánc felsőbb szintjeinek képviselői fogyasztanak (szicíliai ismerőseim esküsznek a tengeri sün pastára), utóbbiakon pedig fontos fejlődésbiológiai modell-szervezetek.
A tengeri sünök kétségtelen, hogy messze nem a legfontosabb és legnépszerűbb állatok (tengerparton ejtőzők szimpátiaindexén valahol egy homokba fúródott üvegcserép és egy különösen éles kődarab között helyezkednek el), de megvan a maguk helye mind a tengerek ökoszisztémáiban, mind a laboratóriumok asztalain. Előbbiekben algával táplákozó primer konzumensek, amelyeket a tápláléklánc felsőbb szintjeinek képviselői fogyasztanak (szicíliai ismerőseim esküsznek a tengeri sün pastára), utóbbiakon pedig fontos fejlődésbiológiai modell-szervezetek.
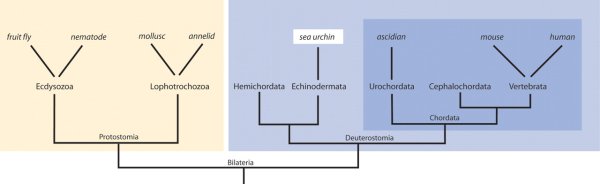
 Különösen alattomos pontja a
Különösen alattomos pontja a