
Ha már a zindex szóbahozott mint releváns hírforrást, akkor álljon itt az általuk is ismertetett cikkhez valóban kapcsolódó post (ui. amire ők hivatkoznak, az egyáltalán nem erről szól, bár kétségtelen, hogy szerepelnek benne az "őssejt" és "spermium" szavak… azóta kivették a hibás hivatkozást)
Annyiban valóban foglalkoztam már a témával, hogy pár napja egy másik kísérletről tettem említést, ahol mind spermiumokat, mind petesejteket próbáltak őssejtekből laboratóriumi körülmények között tenyészteni, s mint említettem egy komoly gond, hogy bár a létrejövő sejtek morfológiailag hasonlítanak az ivarsejtekre, igencsak kérdéses, hogy valóban funkcionálisak-e.
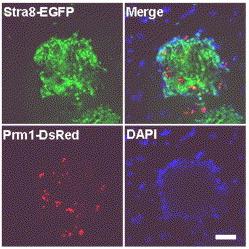
Most német és brit kutatóknak a hímivarsejtek esetében sikerült tovább menniük egy lépéssel és funkcionális spermiumokat létrehozniuk. Az eljárás során retinol-savban (RA) növesztettek olyan embrionális őssejteket, amelyekbe bejuttatak egy, a stra8 gén promóteréhez kötött fluoreszcens fehérjét (EGFP) kódoló génszakaszt. (A stra8 gén jellemzően csak a hímivarsejt-vonalban fejeződik ki, így a promóteréhez kötött EGFP is csak azokban a sejtekben kerül átírásra, amelyek ezirányba kezdtek differenciálódni – értsd az ilyen sejtek megfelelő megvilágítás esetén a GFP miatt zölden fluoreszkálnak.) A GFP pozitív sejteket elkülönítették, és a biztonság miatt egy másik hímivarsejt specifikus gén, a prm1 promóterével össekapcsolt piros fluoreszcens fehérje (DsRed) DNS-ét is beléjük juttatták (bízva abban, hogy azok a sejtek, amelyek mind a zöld, mind a piros fluoreszcens fehérjét expresszálják valóban hímivarsejtek lesznek). Az egyaránt piros-zöld sejteket (a kék színű DAPI a sejtmagok vizualizálására használt) közelebről megnézve, azok valóban expresszálták a spermatogenezishez kapcsolódó géneket, vagyis minden arra mutatott, hogy ezek funkcionális ős-hímivarsejtek.
Már csak az volt hátra, hogy kiderüljön mennyire működőképesek a belőlük létrejövő spermiumok. Ezért előzőleg hímivarsejtjeiktől megfosztott hímek heréibe ültették a sejteket (erre azért volt szükség, mert a here olyan környezetet kínál, amely ideális a hímivarsejtéréshez, s jelenleg nem ismerünk minden faktort, hogy ezt egy lombikban is meg tudjuk ismételni). Néhány hónap múlva a "mostoha" apák heréiben spermium-szerű, DsRed pozitív (vagyis a beültetett sejtekből származó) sejteket leltek. Azonban ezek nem voltak tökéletes hímivarsejtek: mozgékonyáguk egyáltalán meg sem közelítette a normális spermiumokét, s ha mesterséges megtermékenyítésre használták őket, bár születtek kisegerek (ez egyébként nagy fegyvertény, eddig senki sem jutott eddig), azok vagy abnormálisan nagyok vagy abnormálisan kicsik voltak. Ezek pont olyan problémák, amelyek általában klónozás során jelentkeznek és arra az okra vezethetők vissza, hogy ma még csak a felszínét kapargatjuk az epigenetikai változások megértésének.
Az epigenetikai változások (pl. a DNS metilációja) nem okoznak változást a DNS szekvenciájában, de képesek "kijelölni" DNS szakaszokat és befolyásolni később átírásukat ill. átnemírásukat. Az említett "apátlan" kisegerekben (hiszen a kiindulási őssejtek akár nőstényekből is származhattak) is számos DNS-szakasz metilációja volt abnormális, s ez okozhatta a fejlődési rendellenességeiket (egyébként egyikük sem élt többet 5 hónapnál).
Szóval summa-summarum, bár a Szexmisszió árnya kétségtelenül fenyegetőbben tornyosul fölénk, férfitársaim, a veszély még egyelőre nem testközeli ;-).
Nayernia, K, Nolte, J, Michelmann, HW, Lee, HJ, Rathsack, K, Drusenheimer, N, Dev, A, Wulf, G, Ehrmann, IE, Elliott, DJ, Okpanyi, V, Zechner, U, Haaf, T, Meinhardt, A, Engel, W (2006) In Vitro-Differentiated Embryonic Stem Cells Give Rise to Male Gametes that Can Generate Offspring Mice. Dev. Cell. 11: 125-132.

 Darwin pintyei mindmáig a természetes szelekció kinézet (fenotípus) formáló erejének talán legjobb szemléltetői maradtak. Ma már gimis anyag, hogy a Galapagos szigeteken fellelhető magok mérete miként formálta a pinty (Geospiza) fajok csőrének méretét és alakját, s mint az már egyszer
Darwin pintyei mindmáig a természetes szelekció kinézet (fenotípus) formáló erejének talán legjobb szemléltetői maradtak. Ma már gimis anyag, hogy a Galapagos szigeteken fellelhető magok mérete miként formálta a pinty (Geospiza) fajok csőrének méretét és alakját, s mint az már egyszer 

 Hát igen, bennünket is elért a blogok végzete és nevezve lettünk a hvg.hu
Hát igen, bennünket is elért a blogok végzete és nevezve lettünk a hvg.hu 

 Ha már
Ha már