
 Ha már amúgy is a pigmentáció evolúciójáról ejtettem szót, akkor nem maradhat ki egy másik aktuális cikk sem.
Ha már amúgy is a pigmentáció evolúciójáról ejtettem szót, akkor nem maradhat ki egy másik aktuális cikk sem.
Egy kis taxonómiai ugrásra azért szükség van: ezúttal emlősök helyett a jól bejáratott muslicákról lesz szó. A számos Drosophila faj közül jópárra, így a laboratóriumban használt D. melanogaster-re is jellemző a szexuális dimorfizmus, azaz a hímek és a nőstények nem ugyanúgy néznek ki. Ennek egyik leglátványosabb megjelenése a potroh mintázatában fedezhető fel: míg a hímek esetében az utolsó két szelvény teljesen fekete, addig a nőstényeknél ez a két szelvény sem különbözik mintázatában a többi potroh szelvénytől.
A sötét színű pigment termeléséért alapvetően a yellow génről átíródó fehérje felel (a név senkit ne tévesszen meg, Drosophilában aszerint nevezik el a géneket, hogy mi lesz a muslica fenotípusa, ha a gén mutálódik), s mint az a jobboldali ábrán látható (F, ill. G), ez a fehérje épp ott van jelen a lárvákban, ahol később a fekte pigment termelés megindul. Azaz nőstényekben, csak egy vékony csíkban a szegmensek végén, míg hímekben az utolsó két szegmenset teljesen betölti.
A logikus következő kérdés, hogy mi is szabályozza a yellow átíródását, mely gének működése miatt korlátozódik az a fent említett mintázatokra. Itt három gént kell kiemelnünk, az egyik az abd-b, egy homebox gén (ennek domináns mutációjakor pl. a hímek negyedik potroh szegmense is szénfekete lesz), a másik a bric-à-bric (bab), amely két rokon transzkripciós faktort kódol, a harmadik pedig a doublesex (dsx) gén, amelynek van hím és nőstény specifikus variánsa (hogy miért, azt lásd itt). Az abd-b szerepe kettős, mert egyrészt maga is serkenti a yellow átíródását, másrészt pedig kordában tartja a bab gén termékének működését. Utóbbi elnyomja a melanin termelést, pontosabban hímekben csak nyomná, mert ott az ABD-B fehérje kordában tartja ezt a funkciót; nőstényekben ezzel szemben a DSXf fehérje ezt a kordában tartást szünteti meg, így mégsem alakul ki pigmentáció.
Nnna, akkor ennyi molekuláris bevezetés után jöjjön a feketeleves. A muslicák dimorfikus pigmentációjában az is érdekes, hogy bár mint az elején emítettem számos fajban létezik, jópár fajban ugyanakkor hiányzik, ráadásul ezek a fajok sokszor igen közeli rokonságban állnak pigmentált fajokkal. Mivel ez egy viszonylag egyszerű változás és a D. melanogaster mutánsainak vizsgálata azt sugalta, hogy akár egyetlen gén megváltozása is okozhatja ezt, adódott az ötlet, hogy utánna kellene járni, milyen változások is következtek be a különböző fajok között. Az evo-devo (és különösen a gerincetelen hox gének vizsgálatának) egyik nagy öregje, Sean B. Carroll látott neki felgöngyölíteni, mi is történt itt.
Első lépésként a különböző Drosophila fajokban megkeresték a yellow gén átíródását szabályozó szakaszokat (cis-regulatory elements – CRE) és azokat hasonlították össze, úgy, hogy megvizsgálták milyen módon szabályoznák egy D. melanogaster embrióban a gén expresszióját. Mint az alábbi képen is látni, a szexuális dimorfizmust mutató fajokból származó CRE-k, hasonlóan a laboratóriumi muslica sajátjához, különböző mintázatot adtak nőstényekben és hímekben.
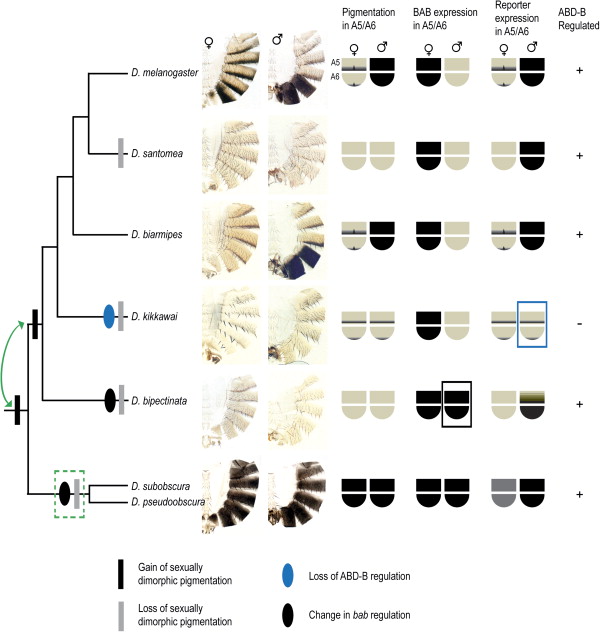
Az ábrán az figyelhető meg, hogy a többi, szexuális dimorfizmust mutató fajból (hogy milyen lesz végül a potroh mintázat, azt az utolsó ábrán lehet látni) – pl. D. biamipes (bia) – származó CRE (a body arra utal, hogy a gén testi expresszióját befolyásoló szakaszokról van szó) különböző módon szabályozzák hímekben és nőstényekben a yellow gén átíródását, míg a dimorfizmust nem mutató, tiszta fehér – D. kikkawai -, vagy fekete – D. pseudoobscura – fajokban a gén vagy sehol, vagy mindenütt expresszálódik. Érdekes, hogy a dimorfizmust nem mutató D. bipectinata (bip) és D. santomea (san) CRE-je is dimorf expressziót hoz létre. Az utolsó sorban levő ábrák azt mutatják, mennyire érzékeny az adott CRE az Abd-B mutációira.
A bevezetőben leírtak fényében talán nem meglepő, hogy a szexuálisan dimorf fajok yellow génjének CRE-jében olyan szakaszokat is leltek, amelyekhez az Abd-B képes kötődni (ez mégis fontos, mert bármilyen sokat is tudunk ma már a Hox-génekről, egyelőre elég rövidke listával rendelkezünk arról, hogy pontosan milyen géneket szabályoznak közvetlenül). Érdekes módon ez a szakasz jelen van a dimorfizmust nem mutató D. bipectinata-ban és D. santomea-ban is, míg a szintén monomorf D. kikkawai-ból hiányzik. Mint arra fény derült, a bipectinata azért lesz monomorf mégis, mert a bab gén valamilyen módon hímekben is kifejeződik, így aztán „felülírja” az Abd-B által kódolt mintázatot. A D. kikkawai CRE-jében pedig két bázispárnyi változás következett be az Abd-B kötőhelyén, s ez elegendő volt ahhoz, hogy a fehérje ne ismerje fel a promotert (ui. ha a labormuslicában is megváltoztatjuk ezt a két bázispárt, ott is megszűnik a yellow gén átíródásának dimorfizmusa). Azaz íme egy kézzel fogható példája annak, miként okozhat fenotipikus változást egy hox gén kötőhelyének a megváltozása!
Az utolsó kérdés az marad, hogy miképpen alakult ki legelőször a dimorfizmus. Első lépésben neutrális mutációk megjelenése során fokozatosan létrejött egy Abd-B kötőhely a drosophilid ős yellow génjének promoter szekvenciájában (a távoli rokonnak számító a D. pseudoobscura és D. subobscura esetében gyenge, de funkcionális Abd-B kötőhelyek vannak, amelyek csak pár bázispárban különböznek a dimorfizmust mutató fajok Abd-B kötőhelyétől) . Hogy aztán mi történt, arra az eddigi eredmények alapján nincs teljesen egyértelmű válasz. Ugyanis az említett távoli rokonok bekavarnak: mindkettőben mindkét nem egész potroha feketében pompázik, de, kicsit váratlanul, a yellow gén szabályozása ennek ellenére enyhe dimorfizmust mutat. Azaz két eset lehetséges, vagy már az említett drosophilid ősben kialakult a dimorfizmus, amely aztán D. pseudoobscura és D. subobscura esetében elvesződött, vagy csak a két vonal szétválásakor (a fenti két faj vs. mindenki más) alakult ki a D. melanogaster-hez is vezető vonalban. Előbbi esetben a bab DSXf függő regulációja is feltehetően létezett az ősben, csak elvesződött a két „outsider” Drosi felé vezető evoúciós úton, míg a második variáció szerint ez a reguláció csak a többi Drosophila felé vezető úton keletkezett, létrehozva a nemtől függő mintázatokat.
(Érdekes még megemlíteni a D. santomea-t is, ahol bár mindkét nem potroha fehér, sem a bab gén szabályozásában, sem a yellow CRE Abd-B kötőhelyében nem volt változás. Itt más ok(ok)ra vezethető vissza a pigmentáció hiánya, de hogy mire, az ma még nem ismert.)
(Hmmm, sajnos gyanítom, hogy túlságosan sok terminus technicus szerepel ebben a postban, de hacsak nem annyit akartam írni, hogy „amerikai kutatók bebizonyították, hogy hox gének szabályozzák egyes Drosophilák szexuális mintázatát”, ezt nem lehetett megkerülni. Ha valaki nagyon elveszett, ígérem, lesz könnyebben emészthető téma is. ;-))
Jeong, S, Rokas, A, Carroll, SB (2006) Regulation of Body Pigmentation by the Abdominal-B Hox Protein and Its Gain and Loss in Drosophila Evolution. Cell 125: 1387-1399.