Nem reprezentatív (főleg az Indexes olimpiai blogok kommentjeinek olvasására kiterjedő) felmérésem szerint, ma már egyre kevesebbeknek ugrik be a Citius, Altius, Fortius hármas az olimpia kapcsán, helyét főként a doppingról való vita véve át. S bár kétségtelen, hogy Pierre de Coubertin báró nem lelkesedne attól, hogy 2008-ban, a Sport Ünnepe idején az a legfontosabb megbeszélni való, hogy most akkor Michael Phelps minden idők legnagyobb sportolója, vagy inkább csak legnagyobb kokszolója, az sem elfelejtendő, hogy ettől a probléma nem újkeletű (NDKs úszólányok, ugye), max csak sokkal többen, sokkal többet beszélnek róla.
Nem reprezentatív (főleg az Indexes olimpiai blogok kommentjeinek olvasására kiterjedő) felmérésem szerint, ma már egyre kevesebbeknek ugrik be a Citius, Altius, Fortius hármas az olimpia kapcsán, helyét főként a doppingról való vita véve át. S bár kétségtelen, hogy Pierre de Coubertin báró nem lelkesedne attól, hogy 2008-ban, a Sport Ünnepe idején az a legfontosabb megbeszélni való, hogy most akkor Michael Phelps minden idők legnagyobb sportolója, vagy inkább csak legnagyobb kokszolója, az sem elfelejtendő, hogy ettől a probléma nem újkeletű (NDKs úszólányok, ugye), max csak sokkal többen, sokkal többet beszélnek róla.
Hogy a doppingolás léte egyeneságú következménye-e annak, hogy a sportban emberek szerepelnek, saját természetük minden hibájával egyetemben, szvsz megérne egy nagyobb lélegzetvételű nekifutást, de a poszt témája szempontjából ez irreleváns. Fogadjuk el a tényt, bármennyire is kellemetlen, dopping volt, van és lesz. Itt és most a doppingolás jövőjéről elmélkednék kicsit, mert már fel-fel sejlik, hogy milyen irányba is mozog ez az „ipar”.
„Géndoppingként” a tiltott előnyszerzés azon (sokak szerint jó eséllyel már talán alkalmazott) formáját aposztrofálják, amikor teljesítményfokozó fehérjék és hormonok szintetikus másolatai helyett, az azokat kódoló gének extra kópiáit juttatja be lokálisan (pl. a mozgás dandárját végző izmokba). Így nemcsak jobb hatásfoka lehet a doppingnak (hiszen a kívánt helyen, többé-kevésbé a kívánt mennyiségben lesz a molekula jelen), de nehezebb detektálni is a folyamatot: például ma a természetes és szintetikus erythropoetin (EPO) közt még kimutatható a különbség, de az extra génkópiákról termelődő extra EPO kémiailag aligha lesz megkülönböztethető a szervezet sajátjától. Igaz, a WADA nem lesz még teljesen tehetetlen: a génbevitel ma csak retrovírusok segítségével lehetséges (ezek biztosítják, hogy az extra gén beépüljön a kromoszómába), márpedig ezen virális vektorok szekvenciája éppoly árulkodó marad arról, hogy valami simlisség történt, mint a puszta tény, hogy a szóbanforgó sportolóban furcsamód több epo génkópia van jelen, mint az elvárható két darab.
Nehezebb a helyzet, ha, teszem azt, adott sportoló sejtjeiből készült sejtvonalban viszik be a szóbanforgó géneket, majd az extra fehérjéket „lefölözik” a tenyészetről, és ezt adják a delikvensnek. A kémiai hasonlóság kvázi ua. jó lesz mint az előző esetben, és mivel a retrovirális inszerció egy titkos laboratórium, titkos Petri-csészéjében esett meg, ellenőr legyen a talpán, aki a dopping-labor lebukása nélkül bizonyítani tudja a turpisságot.
Persze voltak/vannak/lesznek olyan egyedi esetek, amikor nincs szükség semmiféle géndoppingra, az illetőnek természetesen magasabb vörösvérsejt, adrenalin, tesztoszteron, stb. szintje lesz mint társainak. Nincs ebben semmi meglepő, hiszen nem klónok vagyunk hanem emberek. Élőlények, akikre épp úgy élnek a természet és természetes szelekció törvényei, mint bármely más fajra. Amilyen kicsi az esélye két tökéletesen egyforma ujjlenyomatnak, olyan valószínűtlen, hogy két atléta össze teljesítményt meghatározó génje (kódoló és szabályozó szekvenciástúl) bázispárról, bázispárra teljesen megegyezzen. Ez az egyszerű biológiai oka annak, hogy mindig lesznek „tehetségek”, akik kevesebb gyakorlással és odafigyeléssel, jobb eredményeket érnek el dopping nélkül is, mint vért izzadó kollegáik. Hogy a posztot is ihlető, Der Spiegel cikk zona.hu-s átiratát idézzem: „Az átlagos sportolónak egy milliliter vérében három nanogramm tesztoszteron van, a sprinter Tim Montgomerynek is három nanogrammja van, Maurice Greene-nek viszont kilenc. Mit tehet Tim? Nem a dopping igazságtalan, hanem a természet.”
Lehet ijesztő, vagy ígéretes, de mindenképpen valósnak tűnik egy olyan jövőkép, amelyben (főleg azokon a helyeken, ahol csak az eredmény számít, a résztvétel kevésbé), már korai gyermekkorban, géntesztek révén befolyásolódhat, hogy ki, milyen sportba kezdjen, ha vannak élsportoló ambíciói. Már ha ez valamennyire személyes döntése lehet, s nem kell a „nemzet érdekeinek” alávesse magát… (Egyébként, ha már ezt piszkáljuk, vajon mennyire egyeztethető össze a coubertini eszmével a sportolókat testileg, lelkileg tönkretevő edzésprogram?)
(Az illusztráció a The Economist cikkéből származik.)
ui: Gondolom másnak is feltűnt, de az EPO büntetése érdekes logikai dilemmát okoz. Hiszen, ha több hónapot tölt valaki a hegyekben, akkor természetesen is megnöveli a vörösvértesteinek számát, illetve azok oxigén megkötő képességet (részben épp a szervezetben természetesen létrejövő EPO által). Ez, tudtommal, fair, nem büntethető. Ha a tengerszinten él, de elég szerencsés és hozzáférése van speciális, a magaslati viszonyokat szimuláló akklimatizációs szobákhoz, az szintén belefér a pakliba. Ha viszont nincs pénze ilyen kütyükhöz, és akár minimális EPO-t közvetlenül juttat a szervezetébe (pl. annyit amit a szervezete egy magaslati edzőtáborban termelne), az már kiveri a biztosítékot, azzal megsérti a fair-play szabályait.

 Nem ez minden idők legjobb Dawkins filmje, az kétségtelen. Az öreg Richard sok helyen frusztráltnak tűnik – persze nem csoda, hiszen amikor x+1. alkalommal is ugyanazokkal a lejáratott "ellenérvekkel" kell szembesülnöd, amelyek megcáfolására már több könyvet is írtál, nehéz a dolgok pozitív oldalát meglátni. Pedig a bosszús magyarázgatás helyett, (szerintem) egy kis jól irányzott irónia (neagyisten gúny) sokkal hatékonyabb lehet, különösen a képernyőn.
Nem ez minden idők legjobb Dawkins filmje, az kétségtelen. Az öreg Richard sok helyen frusztráltnak tűnik – persze nem csoda, hiszen amikor x+1. alkalommal is ugyanazokkal a lejáratott "ellenérvekkel" kell szembesülnöd, amelyek megcáfolására már több könyvet is írtál, nehéz a dolgok pozitív oldalát meglátni. Pedig a bosszús magyarázgatás helyett, (szerintem) egy kis jól irányzott irónia (neagyisten gúny) sokkal hatékonyabb lehet, különösen a képernyőn.
 Fontos állomásához érkezett, fajunk egykor élt legközelebbi rokonának, a
Fontos állomásához érkezett, fajunk egykor élt legközelebbi rokonának, a  Az autó előtt volt a ló, s ez időben a fehér hátas számított az abszolút státuszszimbólumnak. Nem meglepő hát, hogy a tenyésztők mindent megtettek a privilegizált kevesek ezirányú igényeinek kielégítéséért. E törekvések eredményeként, egymástól függetlenül, több világos ló
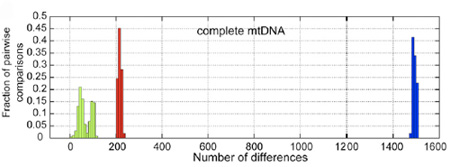
Az autó előtt volt a ló, s ez időben a fehér hátas számított az abszolút státuszszimbólumnak. Nem meglepő hát, hogy a tenyésztők mindent megtettek a privilegizált kevesek ezirányú igényeinek kielégítéséért. E törekvések eredményeként, egymástól függetlenül, több világos ló Több mint 700 ló megvizsgálása után (na és persze a jól haladó
Több mint 700 ló megvizsgálása után (na és persze a jól haladó  "What about all the photos from space that show, beyond a shadow of
"What about all the photos from space that show, beyond a shadow of És
És .jpg) A
A  Mint megannyi más békafaj esetében, az afrikai sarkantyús karmosbéka (Xenopus tropicalis vagy Silurana tropicalis) hímjei, az udvarlás során, hangos éneklés kíséretében csapják a szelet választottjuknak. A hangoskodás fizikailag is megterhelő dolog, olyannyira, hogy a hímek gégeizmai jóval fejlettebbek a nőstényekénél. Ráadásul előbbiek esetében ezek mind gyorsan összehúzódó, ún. kettes típusú
Mint megannyi más békafaj esetében, az afrikai sarkantyús karmosbéka (Xenopus tropicalis vagy Silurana tropicalis) hímjei, az udvarlás során, hangos éneklés kíséretében csapják a szelet választottjuknak. A hangoskodás fizikailag is megterhelő dolog, olyannyira, hogy a hímek gégeizmai jóval fejlettebbek a nőstényekénél. Ráadásul előbbiek esetében ezek mind gyorsan összehúzódó, ún. kettes típusú  A görög mitológia
A görög mitológia  Az állatvilágban közel sem szokatlan, ha egy-egy faj egyedei már embrionális fejlődésük után, de még felnőtté válásuk előtt nagyarányú változáson esnek át (gondoljunk csak a lábat növesztő ebihalakra), a
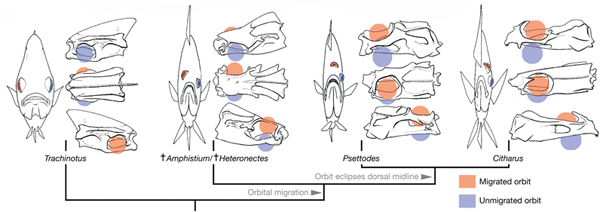
Az állatvilágban közel sem szokatlan, ha egy-egy faj egyedei már embrionális fejlődésük után, de még felnőtté válásuk előtt nagyarányú változáson esnek át (gondoljunk csak a lábat növesztő ebihalakra), a 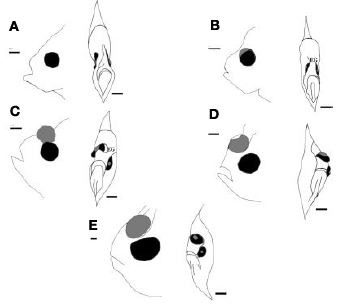 A vándorlás kapcsán azért indokolt a jelenidő, mert ezt a folyamatot minden egyes halnak le kell „játszania” élete során – ugyanis a lepényhalak életük kezdetén még igencsak „normálisak”, s szemük a test két oldalán helyezkedik el (jobb oldali ábra, A). Igaz, ezek a fiatal halak még függőleges testhelyzetben úszkálnak, s ezen viselkedésük is csak a fizikai átalakulásuk, ún. metamorfózisuk (B-E) végére változik meg.
A vándorlás kapcsán azért indokolt a jelenidő, mert ezt a folyamatot minden egyes halnak le kell „játszania” élete során – ugyanis a lepényhalak életük kezdetén még igencsak „normálisak”, s szemük a test két oldalán helyezkedik el (jobb oldali ábra, A). Igaz, ezek a fiatal halak még függőleges testhelyzetben úszkálnak, s ezen viselkedésük is csak a fizikai átalakulásuk, ún. metamorfózisuk (B-E) végére változik meg.