
A fehér orr tünetegyüttesről már volt szó korábban, de egy nagyon fontos kérdés akkor megválaszolatlan maradt: A kézenfekvő magyarázat a megjelenésére nyilván az, hogy a Geomyces destructans nevű gomba Európában egy régen jelenlévő fertőzés, az itteni denevérek már védettek ellene, az utóbbi években került át Amerikába, az ottani denevérek még fogékonyak rá, azért ilyen pusztító a körükben. Viszont egy másik változat is lehetséges, hogy a G. destructans mindkét földrészen jelen volt eddig is, csak annyi történt, hogy Amerikában kialakult egy új, sokkal veszélyesebb törzse. Látszólag nem sok a különbség a két lehetőség közt, de a következményeik óriásiak lehetnek: Ha a G. destructans egy régi kórokozó új törzse, akkor számítani kell rá, hogy előbb-utóbb Európában is megjelenik, itt is elkezdődik majd a denevérpusztulás, viszont ha egy régi kórokozó, ami most került Amerikába, az Európai denevéreket nem veszélyezteti.
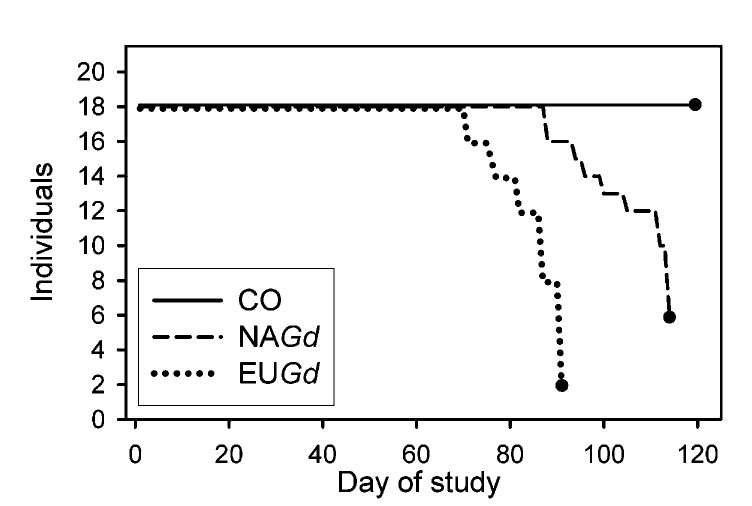 Lisa Warnecke és munkatársai éppen ezt vizsgálták meg, Európából és Amerikából származó G. destructans törzsekkel fertőztek az USÁ -ban gyűjtött denevéreket. Azt tapasztalták, hogy mindkét törzs fertőzése nyomán kialakult a fehér orr tünetegyüttes, sőt, az Európából származó törzs hatására a fertőzött denevérek hamarabb elpusztultak, vélhetőleg ez egy eleve virulensebb törzs. A fertőzött denevérek túlélése látható az ábrán, a folytonos vonal a kontrollcsoport, a két szaggatott vonal a két gombatörzzsel fertőzött két csoport. Jól látható, hogy az Európai G. destructans hamarabb megöli a denevéreket. A következtetés egyértelmű: A G. destructans egy olyan kórokozó, ami itt Európában régóta fertőzi a denevéreinket, Amerikába viszont csak most került át, a mi bőregereink védettek, emberi számítás szerint nem várható a pusztulásuk.
Lisa Warnecke és munkatársai éppen ezt vizsgálták meg, Európából és Amerikából származó G. destructans törzsekkel fertőztek az USÁ -ban gyűjtött denevéreket. Azt tapasztalták, hogy mindkét törzs fertőzése nyomán kialakult a fehér orr tünetegyüttes, sőt, az Európából származó törzs hatására a fertőzött denevérek hamarabb elpusztultak, vélhetőleg ez egy eleve virulensebb törzs. A fertőzött denevérek túlélése látható az ábrán, a folytonos vonal a kontrollcsoport, a két szaggatott vonal a két gombatörzzsel fertőzött két csoport. Jól látható, hogy az Európai G. destructans hamarabb megöli a denevéreket. A következtetés egyértelmű: A G. destructans egy olyan kórokozó, ami itt Európában régóta fertőzi a denevéreinket, Amerikába viszont csak most került át, a mi bőregereink védettek, emberi számítás szerint nem várható a pusztulásuk.
Warnecke L, Turner JM, Bollinger TK, Lorch JM, Misra V, Cryan PM, Wibbelt G, Blehert DS, Willis CK. (2012): Inoculation of bats with European Geomyces destructans supports the novel pathogen hypothesis for the origin of white-nose syndrome. PNAS 109(18):6999-7003.
 Az elevenszülést általában az emlősökre jellemző tulajdonságnak tartjuk, pedig a legkülönbözőbb élőlénycsoportokban alkult ki egymástól függetlenül. Ma egy kevésbé ismert példáról lesz szó, a
Az elevenszülést általában az emlősökre jellemző tulajdonságnak tartjuk, pedig a legkülönbözőbb élőlénycsoportokban alkult ki egymástól függetlenül. Ma egy kevésbé ismert példáról lesz szó, a  Az állatvilágban több a miénktől eltérő rendszer létezik a nemmeghatározásra, ezek egyike a mézelő méh által
Az állatvilágban több a miénktől eltérő rendszer létezik a nemmeghatározásra, ezek egyike a mézelő méh által 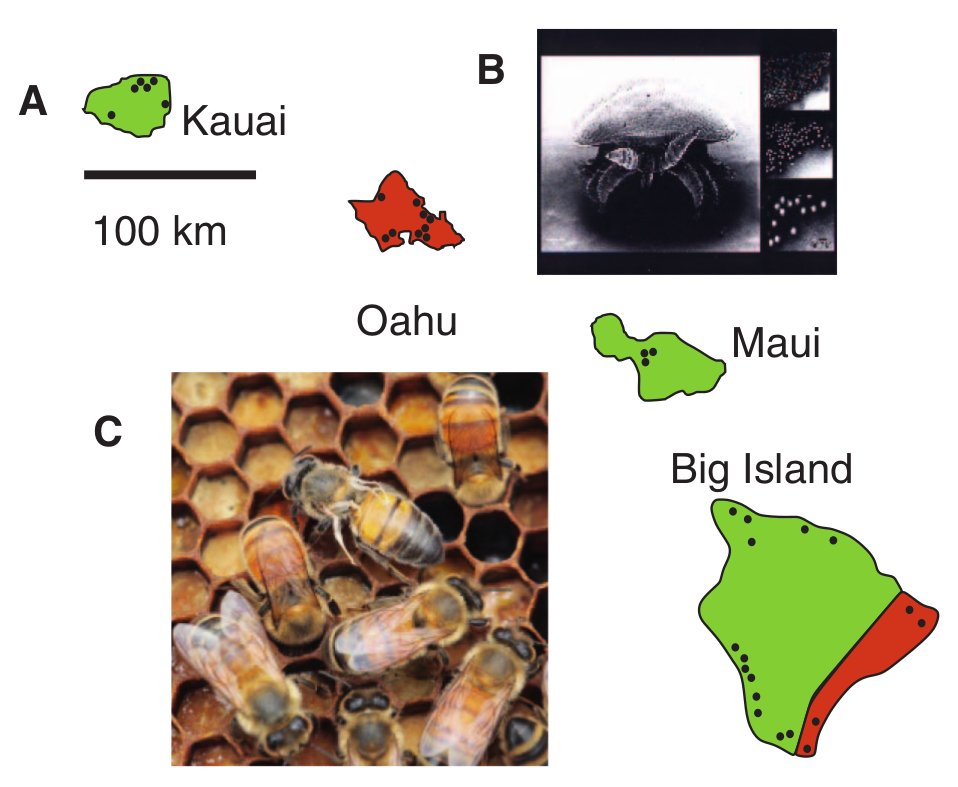 A méhcsalád-összeomlás rejtélyéről volt már szó korábban (
A méhcsalád-összeomlás rejtélyéről volt már szó korábban (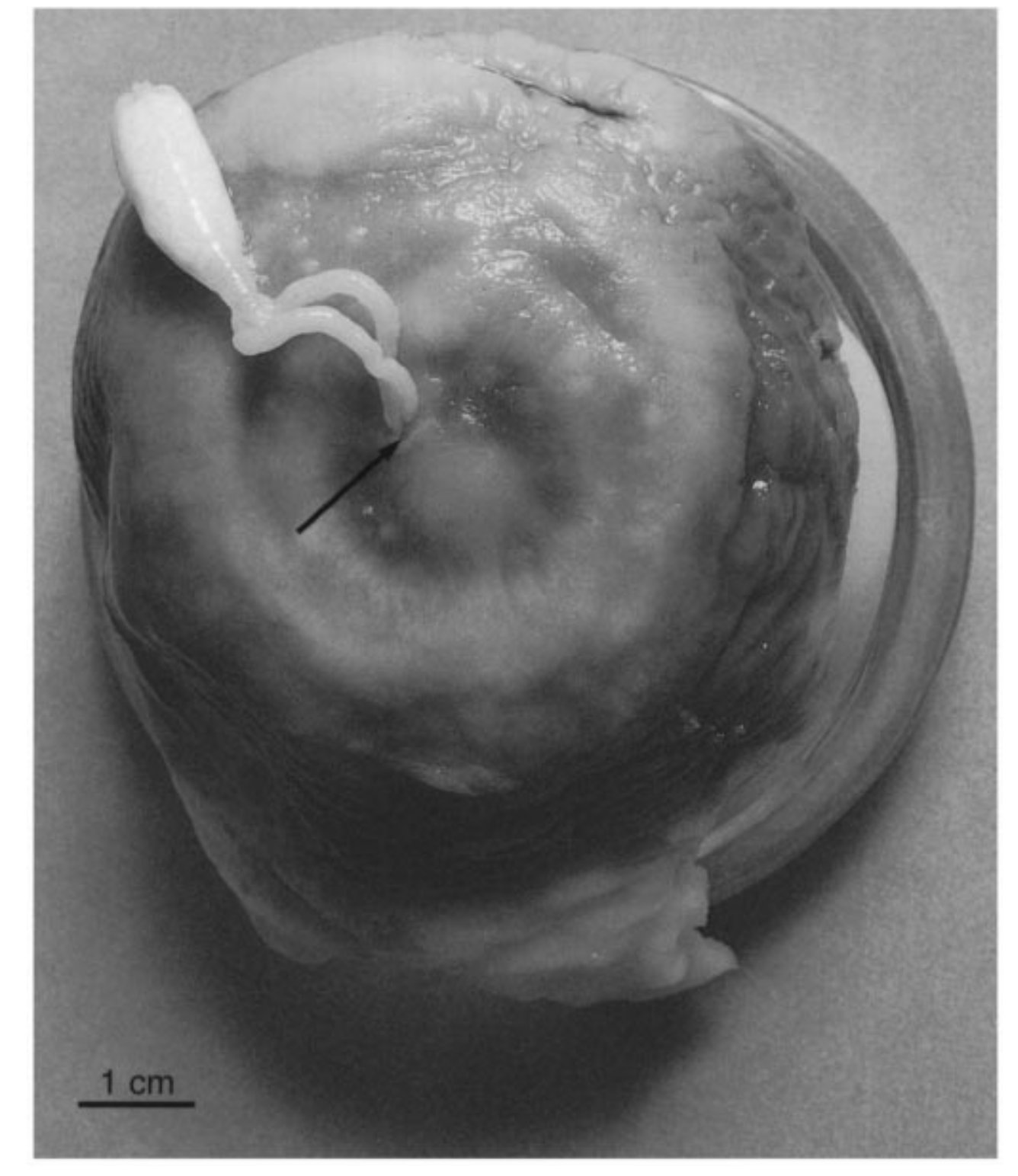 A Grönlandi cápa (Somniosus microcephalus ) egy viszonylag nagy testű, hat méteresnél is hosszabbra növő jószág, amelynek egy sajátos élősködőjéről szól a mai cikk, az Ommatokoita elongata -ról, egy apró, mindössze hat centiméteres rá król. A rák elég kényelmetlen helyre fészkeli be magát, ugyanis a cápa szemgolyójában él, egy külön erre szolgáló szervvel a recehártyára rögzíti magát, úgy, hogy a teste kilóg a vízbe. Ez egy roppant kellemetlen állapot, ráadásul elég gyakori, egy 1959-60 -as felmérés alapján a Grönland mellett kifogott 1505 cápából csak tizenhétben nem találták meg ezt az élősködő rákot (98,9% -os fertőzöttség). Ezen tények alapján joggal merül fel a kérdés, hogy mégis hogyan él túl a Grönlandi cápa, ha ennyire fertőzött a populáció. Az egyik vélemény szerint a rák nem károsítja olyan mértékben a látást, hogy gátolja a gazdát, a másik szerint a faj maga a kihalás szélén táncol.
A Grönlandi cápa (Somniosus microcephalus ) egy viszonylag nagy testű, hat méteresnél is hosszabbra növő jószág, amelynek egy sajátos élősködőjéről szól a mai cikk, az Ommatokoita elongata -ról, egy apró, mindössze hat centiméteres rá król. A rák elég kényelmetlen helyre fészkeli be magát, ugyanis a cápa szemgolyójában él, egy külön erre szolgáló szervvel a recehártyára rögzíti magát, úgy, hogy a teste kilóg a vízbe. Ez egy roppant kellemetlen állapot, ráadásul elég gyakori, egy 1959-60 -as felmérés alapján a Grönland mellett kifogott 1505 cápából csak tizenhétben nem találták meg ezt az élősködő rákot (98,9% -os fertőzöttség). Ezen tények alapján joggal merül fel a kérdés, hogy mégis hogyan él túl a Grönlandi cápa, ha ennyire fertőzött a populáció. Az egyik vélemény szerint a rák nem károsítja olyan mértékben a látást, hogy gátolja a gazdát, a másik szerint a faj maga a kihalás szélén táncol.
 A horizontális génátviteli eseményekről egyre többet tudunk, de engem valahogyan mindig lenyűgöznek. A mai példánk a Hypotenemus hampei nevű rovar, amelyik fontos mezőgazdasági kártevő, ugyanis kizárólag kávén él, a kávétermesztők legnagyobb bosszúságára. Miután a kávébab hatvan százaléka összetett cukrokból áll, a kártevőjének nyilvánvalóan meg kell tudni emészteni ezeket a poliszacharidokat, az ehhez szükséges enzimek pedig a jószág belében kell hogy működjenek. Éppen ezért Ricardo Aru
A horizontális génátviteli eseményekről egyre többet tudunk, de engem valahogyan mindig lenyűgöznek. A mai példánk a Hypotenemus hampei nevű rovar, amelyik fontos mezőgazdasági kártevő, ugyanis kizárólag kávén él, a kávétermesztők legnagyobb bosszúságára. Miután a kávébab hatvan százaléka összetett cukrokból áll, a kártevőjének nyilvánvalóan meg kell tudni emészteni ezeket a poliszacharidokat, az ehhez szükséges enzimek pedig a jószág belében kell hogy működjenek. Éppen ezért Ricardo Aru Mutációkról már volt szó
Mutációkról már volt szó 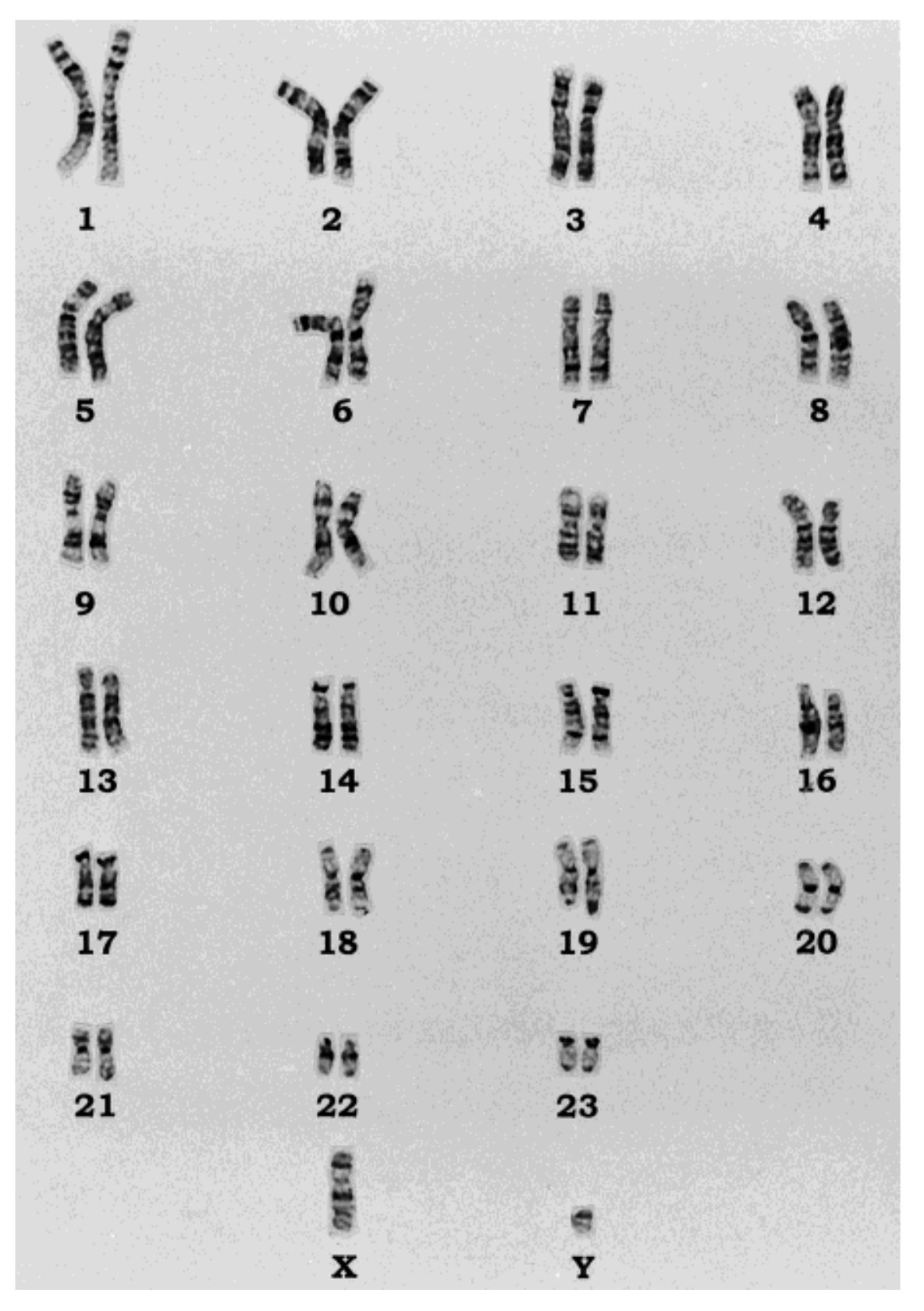 A ma boncolgatott cikk címe kissé szokatlan: „Olivér kromoszómális és mitokonriális DNS elemzése”.
A ma boncolgatott cikk címe kissé szokatlan: „Olivér kromoszómális és mitokonriális DNS elemzése”.