Az egyik gondolat amit időről időre ismételgetni fogok az az, hogy az
evolúció elég szűklátókörű, már olyan értelemben, hogy nem foglalkozik
azzal, hogy milyen változás lehet majd egyszer valamikor előnyös. Ilyen
szempontból sokkal carpe diemesebb a dolog: ami adott pillanatban,
adott körülmények között előnyös az rögzül, ami nem (bármilyen jól is
jöhetne röpke 1 millió év múlva), megy a kukába. Ez a típusú építkezési
logika azzal is jár, hogy olyan változások is rögzülhetnek időnként,
amelyek során egy bizonyos testrész egy olyan funkcióban vállal
hirtelen szerepet amelyhez addig nem volt köze, majd fokozatos
változások révén, a funkció alkalmi hasznos résztvevőjéből
elengedhetetlen összetevőjévé válik. (Ez az úgynevezett koopció jelensége, amelyet a „lecsökkenthetetlen komplexitás” híveinek nem igen sikerül megemészteniük… ;-)).
A koopció egyik szép példája az emlős középfül kialakulása: míg a
hüllőkben egyetlen csontocska (a kengyel megfelelője) játszik szerepet
a hang továbbításban, addig pl. bennünk, három apró csont is található,
a kengyel, a kalapács és az üllő. Ha végignézzük a paleontológiai
leletanyagot jól nyomonkövethető amint a hüllők két állkapocscsontja fokozatosan átalakul a már említett csont-triumvirátus utóbbi két tagjává.
Arról azonban, hogy maga a hüllő középfül hogyan és miért alakult ki, eddig elég keveset tudtunk, ezen változtat azonban a Nature egyik rövid cikke. Martin Brazeau
és főnöke egy lettországi, felső-devonból számazó halszerű kövület
vizsgálata során jöttek rá, hogy egy eddig még le nem írt „átmeneti”
formával állnak szemben: a Panderichthys koponyája ugyanis
egyaránt magán hordoz klasszikus hal- és előbbre mutató jegyeket. A cikk
nem felhőtlenül könnyű olvasmány olyanoknak (pl. jómagam ;-)) akik a
csonttan beszámolókat már sok éve a múlt jótékony feledésébe
süllyesztették, de summa summarum, a lényeg az, hogy a már említett
lény esetében a későbbi közép-fülnek megfelelő helyen egy nagyobb üreg
figyelhető meg, ellentétben a halakkal, viszont hasonlóan a négylábúak
ősének tekintett Acantostegahoz. Ennek a térnek azonban a jelek szerint nem
volt funkciója a hallásban, hanem feltehetően a mai
ráják fecskendőnyílásához hasonlóan, a Panderichthys a
légzéshez, vízáramoltatáshoz használta.
Vagyis a közép-fül
kialakulásában két, időben jól elkülönülő koopciós lépés játszott
fontos szerepet: előszőr egy légzőtérből alakulhatott ki hallótér,
valamikor a szárazföldi gerincesek megjelenésével egyidőben, majd egyes
állkapocscsontok fokozatosan hallócsontokká alakultak.
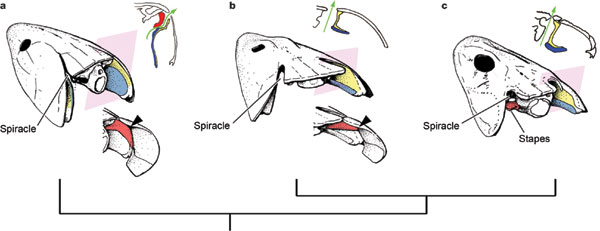
Az ábrán nyomon követhető a középfül terének a kialakulása. (a = Eusthenopteron, b = Panderichthys, c = Acanthostega; a jobb felső sarokban levő ábrák a rószaszín négyzetek magasságában levő keresztmetszeti képet mutatják).
Brazeau MD and Ahlberg PE (2006) Tetrapod-like middle ear architecture in a Devonian fish. Nature 439: 318-321.