
 Hosszú idő óta méltatlanul hanyagoltuk az evodevos posztokat, így most itt az ideje törleszteni ilyen irányú adósságunkat.
Hosszú idő óta méltatlanul hanyagoltuk az evodevos posztokat, így most itt az ideje törleszteni ilyen irányú adósságunkat.
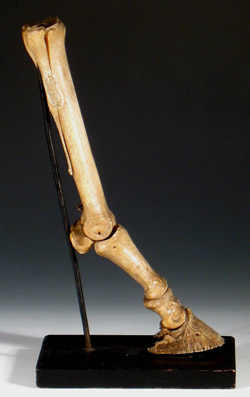
Valamikor a szárazföldi, négylábú gerincesek kialakulásának hajnalán a közös ősben fixálódott az öt ujjú alapvégtag és lényegében a legtöbb élő és kihalt ún. Tetrapoda csoportoban ennek a különböző variációit figyelhetjük meg. Természetesen vannak kivételek (és most eltekintünk a végtagot nem, vagy csak csökevényesen hordozó fajoktól), a legfőképpen a madarak, illetve az emlősök közül a patások, ahol egy és négy között szinte bárhol lehet az ujjak száma.
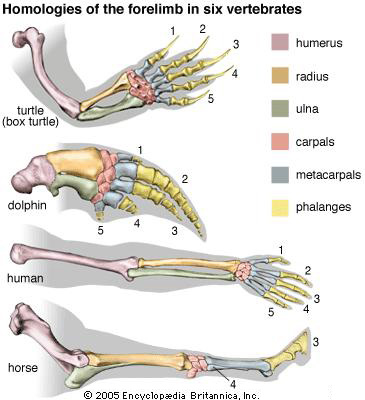
Az emberi csontrendszerhez szokott gondolatvilágunknak furcsa lehet, de a kar- és kéztőcsontok, illetve ujjpercek aránya, illetve egymáshoz viszonyított mérete nagyon eltér a különböző négylábú gerinces fajok között. Míg a mellső végtagjukat úszóként használó teknősök és delfinek esetében az alkar és felkarcsontok sokkal rövidebbek, hogy egy áramvonalasabb úszót hozzanak létre (lásd alább), addig a futásra specializlizálódott lovak esetében az ujjpercek hossza (és vastagsága) már-már az alkarcsontéval (radius) vetekszik. A lovak gyakorlatilag lábujjhegyen járnak, hiszen a lábuk alsó része (lásd jobbra) anatómiailag megegyezik a mi kezünkkel. És hogy a különbség még feltűnőbb legyen, a mi, standard „ötös” kézfelépítésünkkel (szaknyelven pentadactyl végtag) szemben, ők mindössze egyetlen ujjal rendelkeznek.
 Az izgalmas kérdés, természetesen, hogy ez hogyan alakulhatott ki, és erre létezik egyrészt egy evolúciós válasz, másrészt egy fejlődéstani.
Az izgalmas kérdés, természetesen, hogy ez hogyan alakulhatott ki, és erre létezik egyrészt egy evolúciós válasz, másrészt egy fejlődéstani.
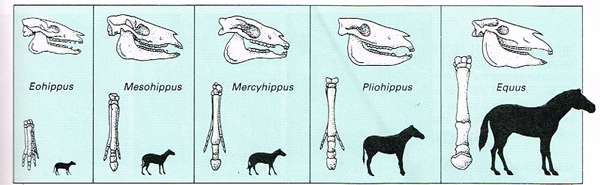
Előbbi már hosszabb ideje ismert volt, a fosszilis anyag gazdagságának köszönhetően. A korai Eohippus kb. egy kutya nagyságú lehetett és még minden végtagján négy ujjal büszkélkedhetett – ezek a mi mutató-, középső-, gyűrűs- és kisujjunknak felelnek meg. (A hüvelykujj különösen érzékeny mindenféle evolúciós változásokra, ennek magyarázata részben a fejlődésében rejlik, lásd ezt a régi posztot, amire majd még sűrűn visszautalok.)
A mai Equus nemzettség felé vezető úton aztán ennek az alapfelépítésnek a további csökevényesedése figyelhető meg a testméret növekedésével párhuzamosan: előbb három, majd egyetlen ujjra redukálódik a végtag felépítése. A kisujj már a Mesohippus esetében eltűnik és ezzel kifejezetten szimmetrikus felépítést nyer a végatg egy robusztus középső, és kevésbé jól fejlett gyűrűs- és mutatóujjal, majd az evolúció során utóbbi kettő fokozatos redukciója figyelhető meg, mígnem teljesen el nem tűnnek (egyébként, ha az embriológiai adatokat nézzük, nem teljesen nyomtalanul, de erre majd hamarosan visszatérünk).
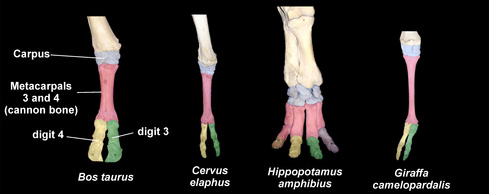
A páratlan ujjú patások mellett (ahova a lovak is tartoznak), természetesen a páros ujjúakat legalább ugyanennyire érinti az ujjszám redukció, ez esetben a négy ujjú vizilovak, és a két-két ujjal rendelkező szarvasmarhák, szarvasok, zsiráfok, vagy tevék mind-mind, a pentadactyl végtag-szerkezet redukciója révén hozták létre saját végtagjaikat (utóbbiak esetében a középső- és gyűrűsujj maradt meg).
 Az evodevo kérdés tehát az lehet ezekben az esetekben, hogy miként alakítható át az a fejlődési program, amely egy ötujjas végtagot hozna létre, úgy hogy a különböző patás csoportokra jellemző szerkezet jöjjön létre.
Az evodevo kérdés tehát az lehet ezekben az esetekben, hogy miként alakítható át az a fejlődési program, amely egy ötujjas végtagot hozna létre, úgy hogy a különböző patás csoportokra jellemző szerkezet jöjjön létre.
Két kézenfekvő megoldás kínálkozik: vagy úgy megváltoztatni az ujjak kialakításához szükséges gradienst, hogy eleve kevesebb ujj jöjjön létre, vagy a fejlődés során fölösleges ujjak fokozatos elsatnyulása következzen be (mintegy haeckeli értelemben másolná így a fejlődési program az evolúciót).
Nos, jelen állás szerint úgy tűnik, különböző fajokban különböző módon következett be a redukció, hol egyik, hol másik előbb részletezett mechanizmust használva, esetleg kettejük kombinációját. A válasz egyik részét Cliff Tabin csoportja adta, akik mongol ugróegereket tanulmányozásától jutottak el a patások ujjredukciójához (mert előbbiek esetében is léteznek olyan fajok, ahol a hátsó lábon csak három ujj van, és az ujjredukció molekuláris mechanizmusa, mint arra fény derült, más fajokra is hasonlónak bizonyult).
A lovak és tevék esetében a jelek arra mutatnak, hogy a programozott sejthalál (apoptózis) biztosítja, hogy a csak egy, illetve két ujj maradjon a fejlődés végére. Az apoptózisnak egyébként igen fontos szerepe van a végtag normális (pentadactyl) fejlődésében is, hiszen ahhoz, hogy az eleinte lapátra emlékeztető végtagkezdeményből kialakulhasson az öt ujj, ahhoz a köztük levő sejtek szabályozott elpusztítására van szükség. Mindez a BMP-jelátviteli útvonal, pontosabban egyik célgénje, az Msx segítségével történik, és ennek a folyamatnak a variálása (pontosabban gátlása) teszi például lehetővé, hogy a kacsák, vagy denevérek esetében az ujjak között megmaradjanak a később fontos élettani funkciót betöltő bőrredők.
A lovak, illetve tevék esetében úgy tűnik, hogy az Msx aktivitásának növekedése következik be a fejlődő végtag két oldalán, aminek hatására felerősődik ezekben a zónákban az apoptózis, és az embrionális fejlődés korábbi fázisaiban még három-, illetve négy ujjas végtag két ujjal kevesebbet hoz majd létre. (Hogy miért nem egy ötujjas alapszabásból indulunk, az egy jó és jelenleg megválaszolatlan kérdés, feltételezhetően itt valami gradiensbeli változás van.)
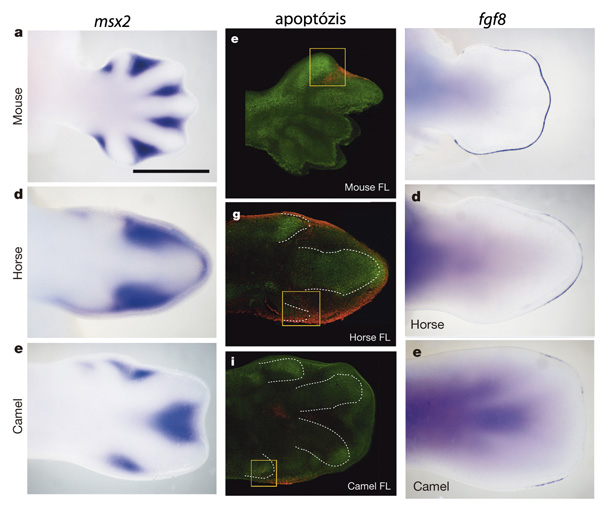
A történet ennyivel nem ér végett, fontos szerepe lesz még a megmaradó ujjak fejlődésében (illetve az elsatnyulásra ítéltek kinemfejlődésében) az ún. apikális ektodermális redőnek (AER) – és ha valaki eddig megspórolta, most itt az ideje, hogy elolvassa a már említett posztot, amiből kiderül, mi is pontosan ez. Az AER a végtagbimbó legszélén található és egy másik jelátviteli útvonal, az FGF ligandjait fejezi ki (pl. fgf8). Funkciója, hogy – az említett ligandok révén – növekedési faktorokat biztosítson, amelyek az alatt növekvő ujjak megnyúlását biztosítják. A patások esetében megfigyelhető, hogy ellentétben pl. egy tipikusan pentadactyl egér-végtaggal, az fgf-ek kifejeződése teljesen szimmetrikus, illetve sokkal kisebb területen figyelhető meg, lényegében mind a lovak, mind a tevék esetében a leendő ujjak felett.
Ez volt tehát az egyik útja az ujjvesztésnek, most lássuk a másikat. Ez, mint azt előre belengettem már inkább az ujjak kialakításában nagyon fontos (harmadik) jelátviteli útvonal, a Shh modulációjával érhető el.
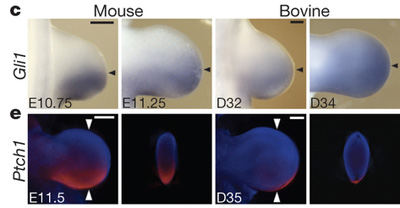 Az Shh-jelátvitel receptora és ligand által nem kötött formában a negatív szabályozója is a Patched (Ptch), amelnyek a kifejeződése a marha embriók végtagjában erőteljesen lecsökkent. Mindez megnövekedett Shh-targetgén aktivitáshoz vezet, például a Gli1 expressziója is lényegesen nagyobb tartományban észlelhető, mint mondjuk egy kontrollként felhasznált egérembrióban.
Az Shh-jelátvitel receptora és ligand által nem kötött formában a negatív szabályozója is a Patched (Ptch), amelnyek a kifejeződése a marha embriók végtagjában erőteljesen lecsökkent. Mindez megnövekedett Shh-targetgén aktivitáshoz vezet, például a Gli1 expressziója is lényegesen nagyobb tartományban észlelhető, mint mondjuk egy kontrollként felhasznált egérembrióban.
Mivel az Shh-aktivitás, pontosabban annak egy gradiens formája az a molekuláris alapmechanizmus, amelyre később az ujjképződés épülni fog, mindez nem meglepő módon ahhoz vezet, hogy megváltozzon – esetünkben lecsökkenjen – az ujjak száma.
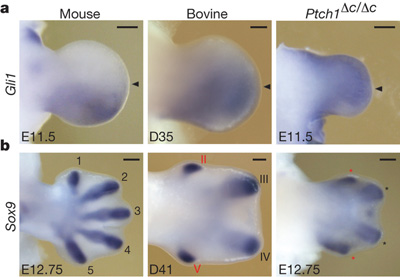 Mindezt persze egyszerűen lehetne, amolyan hasraütéses okoskodásnak is tekinteni, de az egésznek elég jó genetikai alapozást ad, ha kiütjük a Ptch1 gént a fejlődő egérembriük végtagbimbójában.
Mindezt persze egyszerűen lehetne, amolyan hasraütéses okoskodásnak is tekinteni, de az egésznek elég jó genetikai alapozást ad, ha kiütjük a Ptch1 gént a fejlődő egérembriük végtagbimbójában.
Mint a mellékelt ábrán jól látható, ezekben a Ptch1Δc/Δc mutáns végtagokban a Gli1 expressziója hasonlóan kiterjedt lesz egy marhában megfigyelthez, illetve, ha megfesjtük a fejlődő ujjakat, akkor a fejlődés korábbi szakaszaiban ugyanúgy négy darab ujjkezdeményt találunk a mutáns egerekben, mint a vizsgált párosujjú patásban.
De mi okozhatja a Ptch1 lecsökkent expresszióját? A legkézenfekvőbb magyarázat egy szabályozó-mutáció lenne és úgy tűnik, valóban erről van szó.
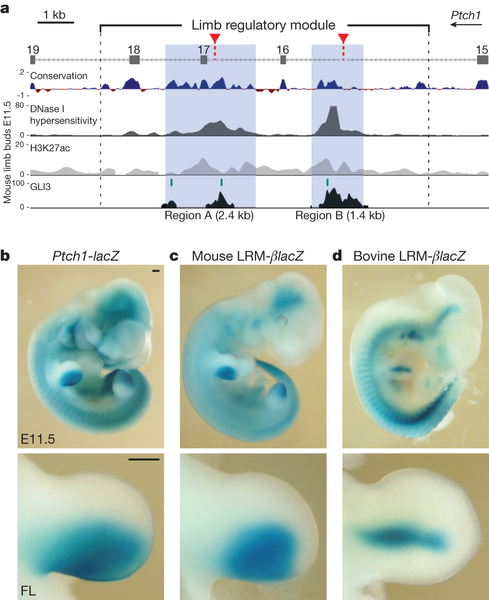 A gén körüli genomi régiókat végigpásztázva fellelhető egy olyan, végtag-specifikus szabályozó modul (limb regulatory module – LRM), amely egy egérben a Ptch1 végtagbimbóban történő kifejeződéséért felel. Magyarán, ha ezt a DNS-szakaszt egérben egy olyan riporter-gén mögé teszem, amelynek a jelenlétét egy egyszerű színreakcióval ki lehet mutatni (pl. lacZ), akkor ezt a színt a végtagbimbó hátsó részében tapasztalom, ugyanott, ahol a Ptch1 egyébként valóban ki is fejeződik.
A gén körüli genomi régiókat végigpásztázva fellelhető egy olyan, végtag-specifikus szabályozó modul (limb regulatory module – LRM), amely egy egérben a Ptch1 végtagbimbóban történő kifejeződéséért felel. Magyarán, ha ezt a DNS-szakaszt egérben egy olyan riporter-gén mögé teszem, amelynek a jelenlétét egy egyszerű színreakcióval ki lehet mutatni (pl. lacZ), akkor ezt a színt a végtagbimbó hátsó részében tapasztalom, ugyanott, ahol a Ptch1 egyébként valóban ki is fejeződik.
Ha azonban nem az egér LRM-et használom a fenti kísérletben, hanem helyette a szarvasmarha LRM-et, lényegesen kisebb területen látom a kék színt, azaz az ebben a régióban megjelenő változások (mutációk miatt lesz a gén lényegesen kevésbé aktív.
Láthatjuk tehát, hogy különösen párosujjú patásokban az apoptózis, a Shh-jelátvitel modulációja, valamint az AER területének szabályozása révén valósul meg az ujjredukció. A szarvasarhák esetében Shh-moduláció egy négy ujjas „alaptervet” hoz létre, és (feltehetőleg) az apoptózis, illetve az AER méretének modulációja lesz az, ami meghatározza mely ujjak maradnak meg és nőnek tovább, míg a többiek fokozatosan elsatnyulnak, elhalnak (pl. a marhák esetében a két szélsőbb ujj az embriogenezis végére rendkívül csökevényes formában, de felismerhető). Fajonként különbözhet azonban a három molekuláris mechanizmus relatív súlya a folyamatban: például tevékben nem változott a Ptch1 expresszió, így itt elsősorban a másik két folyamat hatása érezhető, míg szarvasmarhák és disznók esetében a Shh-jelátvitel (le)szabályozása különösen fontosnak tűnik.
Cooper KL, Sears KE, Uygur A, Maier J, Baczkowski K-S, et al. (2014) Patterning and post-patterning modes of evolutionary digit loss in mammals. Nature doi:10.1038/nature13496
Lopez-Rios J, Duchesne A, Speziale D, Andrey G, Peterseon KA, et al. (2014) Attenuated sensing of SHH by Ptch1 underlies evolution of bovine limbs. Nature doi:10.1038/nature13289