
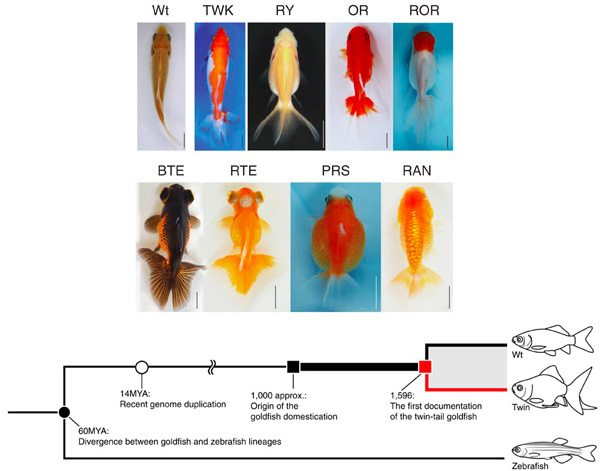
 A bevallottan viccnek szánt kétfarkú kutyával szemben a kétfarkú aranyhalak nagyon is valóságosak: a Ming dinasztia archívumának tanúsága szerint legkésőbb 1596-ban már léteztek a császári udvarban.
A bevallottan viccnek szánt kétfarkú kutyával szemben a kétfarkú aranyhalak nagyon is valóságosak: a Ming dinasztia archívumának tanúsága szerint legkésőbb 1596-ban már léteztek a császári udvarban.
Ekkor az aranyhalakat már közel 600 éve tenyésztették Kínában és mint azt a fajták ma is megfigyelhető formagazdagsága is mutatja, kevés olyan anatómiai korlát volt, amit nem kíséreltek meg több-kevesebb sikerrel áthágni a lelkes domesztikálók.
Akárcsak a kutyák esetében, a domesztikáció lényegében itt is egy hihetetlenül rég zajló és részletes genetikai kísérletként is felfogható, ahol az egyes exotikus fajták fenotípusainak genetikai okait feltárva sok mindent megértünk az állatok „normális” fejlődéséről is.
Az aranyhalak fajtájinak megértésében segítségünkre van az is, hogy egy viszonylag közeli rokonuk a molekuláris fejlődésbiológia modellálata lett: a zebradánióról van szó, amelynek nemcsak a nagyfelbontású genetikai térképe és genom szekvenciája áll a rendelkezésünkre, de az utóbbi évtizedek során számos mutagenezis screen alanya is volt, így nagyon sok, különös mutánssal rendelkezünk, amelyeknek többé-kevsébé értjük a genetikai hátterét is.
A kétfarkú aranyhalakat kutató csoportot is ez segítette a munkájában: első körben azt viszgálták, hogy léteznek-e olyan zebradánió törzsek, amelyek hasonló fenotípust mutatnak, mint a kétfarkú aranyhal.
Mivel kifejlett kétfarkú zebradánió mutáns nincs, a vizsgálatokat kiterjesztették a korai lárvális formákra is (számos olyan mutáció van, ami csak a lárvális fejlődés későbbi szakaszaiban lesz halálos), és ez más sokkal ígéretesebbnek bizonyult.
A pár napos kétfarkú aranyhallárvák szintén többszörös farokfodorral rendelkeznek, és ez már nagyon is emlékeztet egy igencsak ismert zebradánió mutánsra, a chordino-ra (amit néha csak dino-ként aposztrofálnak).
A chordino mutáció a halak chordin (chd) génjét érinti, amelynek, mint minden korán kifejeződő BMP-jelátvitel antagonistának, kulcsfontossága van az embrionális hát-hasi (szakzsargonban dorzoventrális) tengely kialakításában, pontosabban a háti, azaz dorzális oldal kialakításában. (Erről kicsit részletesebben itt írtam, anno.) Fontos itt megjegyezni, hogy a dorzális oldalon kifejeződő gének a leendő idegrendszer és a test elülső részén kialakuló szövetek indukciójában játszanak szerepet, a ventrális gének pedig a kültakaró, illetve a hátsó testtájon kialakuló szövetekért felelősek.
 Mivel a dorzális és ventrális szövetek egymás rovására alakulnak ki (alapesetben egyensúlyban), a hibás dorzális géneket hordozó zebradánió mutánsok, így a chordino is, több ventrális szövetet hordoz: például több vérsejt és több farokredő alakul ki – és ezzel vissza is értünk a kétfarkú aranyhalakhoz, ahol pont ezt lehet megfigyelni.
Mivel a dorzális és ventrális szövetek egymás rovására alakulnak ki (alapesetben egyensúlyban), a hibás dorzális géneket hordozó zebradánió mutánsok, így a chordino is, több ventrális szövetet hordoz: például több vérsejt és több farokredő alakul ki – és ezzel vissza is értünk a kétfarkú aranyhalakhoz, ahol pont ezt lehet megfigyelni.
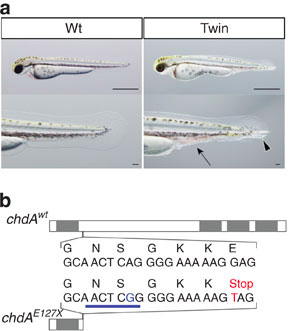
Adta tehát magát az ötlet, megnézni, hogy mi is van az aranyhalak chd génjével. Mint kiderült, a chdA génben valóban van ún. frameshift mutáció (chdAE127X), ami eltolja a fehérje leolvasási keretét és egy korai Stop kodont hoz létre. Az így kialakuló fehérje működésképtelen. A kétfarkú fenotípus és a mutáció közti ok-okozati viszonyt erősíti, hogy az összes kétfarkú fajtában homozigóta formában van jelen a chdAE127X mutáció, míg a „vad fenotípusú”, normál aranyhalakban nem, illetve, ha a nem-mutáns chd génről készült mRNS-t injektáljuk kétfarkú aranyhalak embrióiba (amelyek egyébként szüleik fenotípusát örökölnék), azok nem fejlődnek kétfarkúvá.
Egy kérdés azonban nyitva maradt: a chordino zebrahalak 5-7 napos korukban elpusztulnak, pont a dorzális szöveteik elsatnyulása miatt. Hogy lehet vajon, hogy az aranyhalak ennyire strammok?
A megoldáshoz egy kicsit vissza kell utalnom az aranyhal-evolúciót ábrázoló egyik felsőbb ábrára. Ezen jól látszik, hogy uszkve 14 millió évvel ezelőtt, vagyis lényeges a zebradániók felé vezető vonaltól való elválás után, az aranyhalak ősei egy genomduplikáción mentek keresztül. Az ilyesmi nem idegen a halaktól, már a két halfaj közös ősében is volt egy genom duplikáció, így a csontos halakban nagyon sok génből, amiből belőlünk egy van, kettő (vagy több) is létezik.
Mindenesetre az aranyhalak genom duplikációja ahhoz vezetett, hogy két chd génjük maradt, ugyanis a chdA mellett található egy chdB is. A kettő szekvenciájában (s így feltételezett funkciójában) hasonló, de kifejeződési tartományuk eltérő, bár átfedő: a chdA, hasonlóan az egyszem zebradánió chd-hoz, az úgynevezett gasztruláció során a leendő embrionális tengelyben, valamint kétoldal a gastrula dorzális szélén fejeződik ki. (Ha ez kínaiul hangzik, ne add fel, itt egy ábrasorozat, miként is történik a zebradánió fejlődése – a nagy bumszli, amit körbenőnek a sejtek, a szikanyag, ami a fejlődő embriót, majd a lárvát táplálja, mindaddig, míg az el nem kezd enni.) A chdB ezzel sezmben csak a dorzális tengelyben található meg (lásd alábbi ábrán).
Ez azt jelenti, hogy mutáns chdA funkciója helyettesíthető a dorzális tengelyben a chdB által, viszont a dorsolaterális oldalon nem. Ezért lehet az, hogy számos ventrális gén expressziója megnő (alább a sizzled példája látható), s így az általuk meghatározott szövetekből is több lesz, ugyanakkor az idegrendszer mérete többé-kevésbé változatlan marad (ezt jelzi a korx20 nevű nyúltvelői markergén expressziós doménjének kvázi változatlansága).
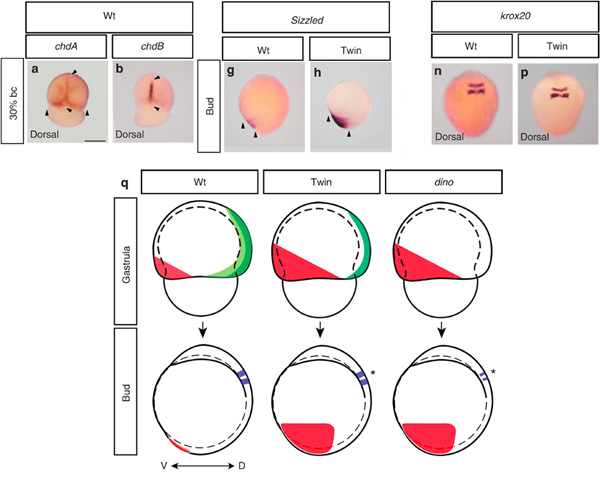
Ez látható a fenti ábrán is (oldalnézetből nézzük az embriót – a ventrális szövetek pirossal láthatók, a chdA és -B domének világos, illetve sötét zölddel), ahol a chordino-val szembeni különbség, az idegszövet méretének relatív változatlansága is jól látható.
Vagyis az egész eset jól mutatja, hogy genom-duplikáció után, korábban sok funkciót ellátó (pleiotróp) gének másolatai önálló evolúciós úton járnak, szubfunkcionalizálódhatnak, és gyakran olyan változásokat tesznek lehetővé, amelyek a duplikáció miatt, az anya-gén sokrétű feladati okán halálosak lettek volna.
Érdekes módon, amikor egyszer már volt ilyesmiről szó, akkor szintén a zebradánió apropóján történt: a spiegeldanio mutáns egy FGF-jelátviteli útvonal receptorában (fgfr1a) hordoz mutációt, hasonlóan a tükörpontyhoz, ami csak azért nem letális számára, mert a szóbanfrogó gén sok funkcióját ellátja az fgfr1b nevű paralógja.
Abe G, Lee SH, Chang M, Liu SC, Tsai HY, Ota KG (2014) The origin of the bifurcated axial skeletal system in the twin-tail goldfish. Nat Commun 5: 3360.