
 Akik járatosak a témában, biztos felfigyeltek rá, hogy mikor a múlt héten az emberi sokféleség eredetét (pontosabban amit erről ma tudunk/gondolunk) igyekeztem zanzásítani, talán kicsit gálánsan csak az egyik létező elméletre koncentráltam, az ún. „out of Africa”, azaz „ki Afrikából” hipotézisre.
Akik járatosak a témában, biztos felfigyeltek rá, hogy mikor a múlt héten az emberi sokféleség eredetét (pontosabban amit erről ma tudunk/gondolunk) igyekeztem zanzásítani, talán kicsit gálánsan csak az egyik létező elméletre koncentráltam, az ún. „out of Africa”, azaz „ki Afrikából” hipotézisre.
Ennek két főbb oka van: egyrészt a genetikai vizsgálatok (amelykről a poszt leginkább szólt) egyértelműen ezt támasztják alá, másrészt (az előbbiek miatt) ez az elmélet ma már nagyságrendileg elfogadottabb a konkurensénél, az ún. „multiregionális” elméletnél. Utóbbi neve kicsit megtévesztő, hiszen alapvetően e szerint is Afrikából származik az emberiség, de a gondolatmenet hívei szerint nem a Homo sapiens ~60,000 évvel ezelőtti kivándorlása volt a kulcsmozzanat, hanem az ennél lényegesen hamarabb történt H. erectus expanzió.
Ennek a félmillió évvel ezelőtt bekövetkezett vándorlásnak számos nyoma van és a jelenleg legelterjedtebb nézet szerint mind a neandervölgyi ember, mind (ha létezett) a Flores szigeti hobbit ennek nyomán keletkezett. Csak míg a „ki Afrikából” hívek szerint a H. erectus Afrikán belüli evolúciója hozta végül létre a modern embert, addig a „multiregionalisták” szerint (akik elsősorban a fossziliákra bazirozzák gondolatmenetüket) az egyes kontinenseken élő emberi rasszok párhuzamosan jöttek létre a lokális H. erectus populációkból (persze azért jelentős keveredéssel, különben aligha maradhatott volna fenn egyetlen fajként az emberiség).
A „multiregionális” elmélet azonban nehezen (vagy úgy sem) tud megbirkózni a molekuláris eredményekkel, hiszen azok nemcsak az emberi populációk kontinuitását igazolták, de rendre azt is mutatták, hogy Afrikától távolodva egyre csökken a genetikai variabilitás. Utóbbi (a „ki Afrikából” elmélet szerint) azért van így, mert ahogy az emberiség lassan, de biztosan egyre távolodott az „őshazától”, az új területeket kolonizálóknak mindig csak egy része (ill. az ő leszármazottaik) ment még tovább, ami óhatatlanul is azt okozta, hogy ebből a vándorló kis közösségből hiányzó gén variánsok, azaz allélok, nem terjedhettek el az új területeket belakó populációkban (és ezért csökkent a diverzitás).
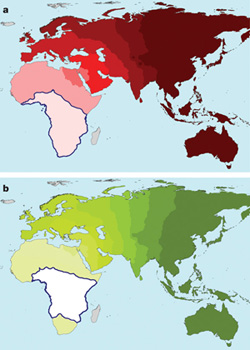
Most azonban úgy tűnik, hogy egy, a Nature-ben megjelent cikk, pontot tesz a vita végére. Egy japán professzor korábban nem publikált adatai (egy masszív, több mint 6200 koponyát felölelő craniometriai adathalmazról van szó!) szintén a „ki az Afrikából” hipotézist támasztják alá. Ugyanis ezek a mérések, a genetikai vizsgálatokhoz hasonlóan, azt mutatják, hogy Afrikától távolodva csökken az emberi koponyák kinézetbeli variabilitása. És bár nyilván a koponya alak nem független a génektől, mégis jó látni, hogy a pusztán DNS vizsgálatokon alapuló eredmények (a zöld színű térkép) és a csonttani mérések (piros térkép) milyen szépen egybeesenk. (A világosabb szín nagyobb változatosságot jelöl, a kékkel körbekeretezett régió pedig az emberiség kialakulásának feltételezett helyét mutatja. Ha a „multiregionalitás” lett volna a nyerő, az egyes kontinensek közepén kellene lássuk a világos foltokat, és köztük a sötétebb sávokat.)
Manica A, Amos W, Balloux F, Hanihara T (2007) The effect of ancient population bottlenecks on human phenotypic variation. Nature 448: 346-348.


 Szűk egy éve
Szűk egy éve  Ha már néhány napja egy ismert ökológiai szabályt igazoló megfigyeléről
Ha már néhány napja egy ismert ökológiai szabályt igazoló megfigyeléről  Ha véginézünk a
Ha véginézünk a  A szigetek élővilága mindig különleges, gondoljunk csak Új Zéland egészen egyedi madár faunájára vagy Galapagos pintyeire és
A szigetek élővilága mindig különleges, gondoljunk csak Új Zéland egészen egyedi madár faunájára vagy Galapagos pintyeire és 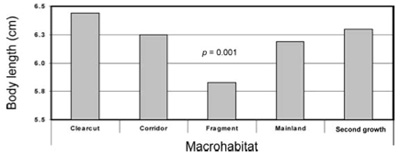 A Washington államban végzett kutatás végén a kutatók, több rágcsáló- (egér- ill. pocok-) fajt vizsgálva arra jutottak, hogy azok mérete az utóbbi mintegy három évtized alatt (ennyi a max. kora a vizsgált élőhely-töredékeknek) szignifikánsan csökkent más élőhelyen leledző rokonaikhoz képest. Mivel az egerek amúgy is igen sikeresek
A Washington államban végzett kutatás végén a kutatók, több rágcsáló- (egér- ill. pocok-) fajt vizsgálva arra jutottak, hogy azok mérete az utóbbi mintegy három évtized alatt (ennyi a max. kora a vizsgált élőhely-töredékeknek) szignifikánsan csökkent más élőhelyen leledző rokonaikhoz képest. Mivel az egerek amúgy is igen sikeresek 