
 Az Enigma rejtélyének megfejtése, és a Turing-teszt megalkotása után Alan Turing új kihívásokat keresett és azokat végül a biológiában találta meg. Egész pontosan a biológiai minták matematikai leírása és képződésük feltételezett mechanizmusa kezdte izgatni zseniális elméjét és ennek a nyomát őrzi legendás 1952-es cikke, amelyben (egyebek mellett) megalkotta a foltszerű mintázatok leírására és modellezésére mindmáig leginkább használt „reakció-diffúzió mechanizmust”.
Az Enigma rejtélyének megfejtése, és a Turing-teszt megalkotása után Alan Turing új kihívásokat keresett és azokat végül a biológiában találta meg. Egész pontosan a biológiai minták matematikai leírása és képződésük feltételezett mechanizmusa kezdte izgatni zseniális elméjét és ennek a nyomát őrzi legendás 1952-es cikke, amelyben (egyebek mellett) megalkotta a foltszerű mintázatok leírására és modellezésére mindmáig leginkább használt „reakció-diffúzió mechanizmust”.
Turing zsenialtása már rögtön ott tettenérhető, hogy ráérzett, a látszólag random vonalak és foltok összessége jól megfogalmazható matematikai törvényszerűségeknek engedelmeskedik, amelyeknél elég néhány paraméter megváltoztatása és egészen különböző mintázatok leírását érhetjük el segítségükkel.
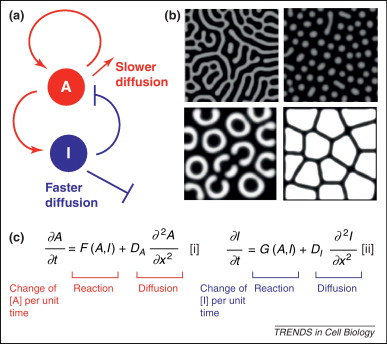 A szabály lényege, hogy ha van egy A aktivátor molekulám, ami pl. pigmentkialakulást okoz (de kültakarón kívül, mindez igaz minde más, molekuláris szinten detektálható mintára). Az A rendelkezik egy adott diffúziós konstanssal, meg persze azzal a tulajdonsággal, hogy aktiválja önmagát és I-t is, saját inhibitorát, jellgezetes pozitív illetve negatív feed-back köröket hozva létre. I szintén rendelkezik egy diffúziós konstanssal, ami történetesen nagyobb, mint az A megfelelő értéke. Ha mindez megvan, akkor minden adott a gyönyörű mintázatok kialakulásához.
A szabály lényege, hogy ha van egy A aktivátor molekulám, ami pl. pigmentkialakulást okoz (de kültakarón kívül, mindez igaz minde más, molekuláris szinten detektálható mintára). Az A rendelkezik egy adott diffúziós konstanssal, meg persze azzal a tulajdonsággal, hogy aktiválja önmagát és I-t is, saját inhibitorát, jellgezetes pozitív illetve negatív feed-back köröket hozva létre. I szintén rendelkezik egy diffúziós konstanssal, ami történetesen nagyobb, mint az A megfelelő értéke. Ha mindez megvan, akkor minden adott a gyönyörű mintázatok kialakulásához.
(A mellékelt ábra képletei ne ijesszenek meg senkit, tényleg nagyon könnyen megemészthetők. F(A,I) és G(A,I) a két molekula közti kölcsönhatást írja le, ami A szemszögéből nézve gátlás, I esetében pedig aktiválás, a DA és DI a két diffúziós konstans.)
A kiindulási állapot általában csak stochasztikus különbségeket rejt az aktivátor koncentrációjában, de ezek elegendők lesznek, hogy a pozitív feed-back segítségével az aktivátor ezeken a helyeken feldúsuljon. Ugyanakkor az inhibitor megjelenése és gyors diffúziója biztosítja, hogy ezek a feldúsulások csak lokálisak maradjanak és ne az egész szövet ússzon az aktivátorban. (Az alábbi ábrán A-ból P, I-ből pedig S lett, de egyébként minden ua..)
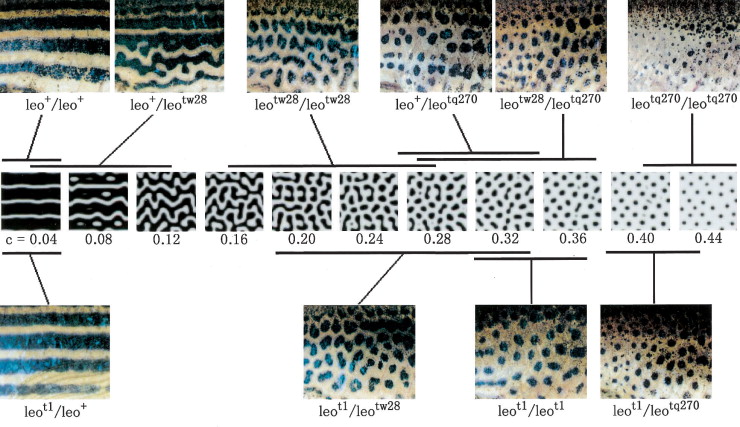
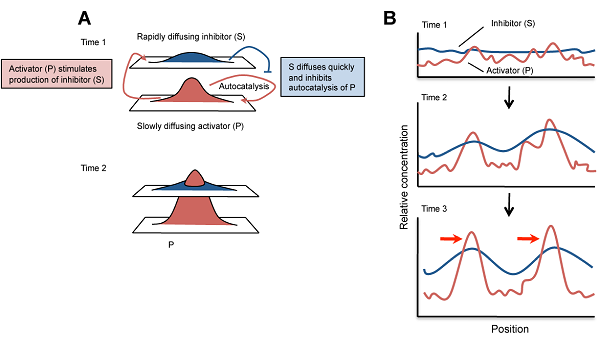 Ennyi a lényeg, a kezdeti paraméterek beállítása aztán majd ontja a minta-kavalkádokat. És ami még jobb: minden jel arra mutat, hogy ezek nagyon is jó leírói lesznek a valóságnak. Az alábbi ábrán egy pigmentáció mutáns zebradánió törzs (leopard) különböző erősségű mutáns alléjainak hordozóit láthatjuk összevetve egy szimulált mintasorozattal, ahol az aktivátor alap-szintétisének szintje – ezt fejezi ki a c – változik csak.
Ennyi a lényeg, a kezdeti paraméterek beállítása aztán majd ontja a minta-kavalkádokat. És ami még jobb: minden jel arra mutat, hogy ezek nagyon is jó leírói lesznek a valóságnak. Az alábbi ábrán egy pigmentáció mutáns zebradánió törzs (leopard) különböző erősségű mutáns alléjainak hordozóit láthatjuk összevetve egy szimulált mintasorozattal, ahol az aktivátor alap-szintétisének szintje – ezt fejezi ki a c – változik csak.
Persze nem a kültakaró mintája az egyetlen példa: a főbb jelátviteli útvonalak – Wnt, Nodal – egyaránt ezt a logikát használják önszabályozásra, ahogy még néhány más fejlődésbiológiai folyamat működése is ebbe az irányba mutat, de ezekről majd később.
Még több matematika a fejlődésbiológiában.
(A fiatal császárhal képe innen származik. Enigma rajongóknak pedig ez kötelező darab.)
A.M. Turing (1952) The chemical basis of morphogenesis. Phil. Trans. R. Soc., B237: 37–72.
Torii KU (2012) Two-dimensional spatial patterning in developmental systems. Trends in Cell Bio 22(8): 438–446.
Asai R, Taguchi E, Kume Y, Saito M, Kondo S (1999) Zebrafish leopard gene as a component of the putative reaction-diffusion system. Mech Dev 89(1-2): 87-92.
Koszi, nagyon erdekes ez is!
A foltok kialakulasat a magyarazat alapjan ertem, vagy legalabb is sejtem. A szabalyos csikok, amik a vad tipusu zebra danion vannak, azok is magyarazhatoak igy?
@fordulo_bogyo: @fordulo_bogyo: igen (sőt az igazi zebrákéi is). kicsit talán baj is: ami túl sok mindent magyaráz, az nem magyaráz semmit:).
googlizz rá a „reaction-diffusion”-re (képkeresővel). amúgy ilyen mintákat létre lehet hozni petricsészében, a megfelelő anyagok összekeverésével. mégjobbak a kétkomponensű fémek olvadékaiban fagyás során kialakuló dendritek. ezt is lehet hasonlóképpen modellezni (phase field theory), és ebben egy magyar kutatócsoport –Gránásyék– is nagyon jó.
nézd végig ezeket a képeket, sokkal-sokkal menőbb, mint a foltok:):
http://www.szfki.hu/~grana/crystal.html
kicsit hasonló még a frontokhoz, szolitonokhoz (csak ott a nemlinearitás és a diszperzió összjátéka hoz ki egy nagyon stabil megoldást, és persze az időben másodrendű (hullámegyenlet))
dolphin: engem az érdekelne, h az egyenlet paramétereit (konkrétan ebben az esetben) és a kölcsönhatási tagokat honnan vették? megmérték (a diffúziót tippem szerint nagyon könnyű mérni)? egyáltalán a kölcsönhatás konkrét alakjára mennyire érzékeny a rendszer (tipp: nagyon)? mert akkor pláne érdekelne, h mi a pontos alakja, és azt honnan vették (tipp: a legegyszerűbb fvényt keresték, ami a megfelelő mintát hozza:)), el nem tudom képzelni, h ezt hogy lehetne rendesen mérni.
@ppp2:
„egyáltalán a kölcsönhatás konkrét alakjára mennyire érzékeny a rendszer (tipp: nagyon)? „
Eléggé.
„el nem tudom képzelni, h ezt hogy lehetne rendesen mérni.”
Nem pontosan ezt mérik, de érdekes lehet számodra.
angel.elte.hu/cellmotility/
” kiindulási állapot általában csak stochasztikus különbségeket rejt az aktivátor koncentrációjában, de ezek elegendők lesznek, hogy a pozitív feed-back segítségével az aktivátor ezeken a helyeken feldúsuljon. Ugyanakkor az inhibitor megjelenése és gyors diffúziója biztosítja, hogy ezek a feldúsulások csak lokálisak maradjanak és ne az egész szövet ússzon az aktivátorban.”
Azért ez nem teljesen így van. Egy reakció diffúzió egyenletrendszerben az időtől nem függő stabil megoldás körülik kis amplitudójú zavarok viselkedése attól függ, hogy milyenek az egyenlet paraméterei és milyenek a geometriai viszonyokt. Bizonyos feltételek mentén lesz oszcilláció, mintázatképződés stb, máskor pedig nem. Ráadásul a kezdeti feltételekben „csak stochasztikus különbségek” nagyon sok mindent befolyásolnak. Konkrétan erős korrelálatlan térbeli zaj szerintem semmilyen RD rendszerben nem hoz létre oszcillációt.
Az ELTE-n egyébként vannak a dolognak kutatói mind a Kémia, mind a Fizika, mind a Matematikai Intézetben. Kurzusokat is szoktak tartani.
Természetesen a titeket érdeklő témában (matekdevo?) is van magyar kutató, Forgács Gábor
forgacslab.missouri.edu/
Könyve is van
http://www.amazon.com/Biological-Physics-Developing-Embryo-Forgacs/dp/0521783372
.
@Apeiron: köszönöm. kicsit utána mentem, ezt érdemes megnézni:
intl-rsfs.royalsocietypublishing.org/content/2/4/487.full
a matek az elején senkit ne riasszon el, ezek az egyenletek a posztban szereplő modellekre példák (1.1 is, ha a diffúziómátrix diagonális, u kétdimenziós és f-et megfelelően választjuk). megnézeget olyan dolgokat is, mint a kezdeti feltételek hatása, finoman utal a perem fontosságára, de nem megy bele részletesen (az nagyon nem meglepő, h a peremfeltételekkel való játszadozásnak drámai hatása van a mintázatra), a zaj és a domain növekedésének hatása.
megnéztem, a Farkas Miklós „Dynamical Models in Biology” könyvében foglalkozik kicsit a kérdéssel (5.2.es alfejezet), nagyon egyszerű matekkal. hivatkozik egy cikkre, amit nem tudtam elérni: Thomas, D. (1975). Artificial enzyme membranes, transport, memory and oscillatory phenomena, in Analysis and Control of Immobilized Enzyme Systems,
Berlin: Springer-Verlag, 115-150.
ebben állítólag empirikus adatokból vezeti le az egyenleteket. ez érdekelne. ha valaki meg tudja szerezni, szóljon.
kapcsolódik még: Meinhardt: Models of Biological Pattern Formation, letöltöttem, de még nem olvastam bele.
@ppp2: BME könyvtár, raktári jelzet 246.128
aleph.omikk.bme.hu/F/VP7IQU9T4SD2BRDSYIVDXD24V9ANU32IMC1BCVAEU9N39QGK75-05522?func=find-b&request=Analysis+and+control+of+immobilized+enzyme+systems&find_code=WRD&x=0&y=0&filter_code_1=WLN&filter_request_1=&filter_code_2=WYR&filter_request_2=&filter_code_3=WYR&filter_request_3=
@Apeiron: köszönöm.
Azzal egyetértek, hogy Turing zseniális, és ez a jelenség is az, de maga a gondolat szerintem trivialitás.