
 Ha Tubifex-ről esik szó, akkor jó eséllyel mindenkinek csak az izgő-mozgó, gorgófő-szerű halkaja csomó ugrik be és legkevésbé sem az, hogy ez a féregfaj is a fejlődésbiológia egyik modellállata.
Ha Tubifex-ről esik szó, akkor jó eséllyel mindenkinek csak az izgő-mozgó, gorgófő-szerű halkaja csomó ugrik be és legkevésbé sem az, hogy ez a féregfaj is a fejlődésbiológia egyik modellállata.
Pedig ez a Kevéssörtéjű férgek csoportjába tartozó állat már hosszú évtizedek óta kitartó figyelmet kapott pár labor részéről, így fejlődéséről is viszonylag sokat tudunk. Standard spirális osztódással indul a történet (akárcsak a pár napja megénekelet csigák esetében), de például a közeli rokon Többsertéjű férgektől eltérően nagyon hamar feltűnik, hogy az első osztódások igencsak aszimmetrikusak, és két igen különböző méretű sejtet hoznak létre. Általában elmondható, hogy a nagyobb sejtből a kifejlett (felnőtt) állat szervez fognak kialakulni, míg a kisebb sejteknek csak kevésbé jól definiálható, obskurus szerep jut. Mindez azért van így, mert az említett Többsertéjűekkel ellentétben itt nem történik meg jellegzetes alakú lárvák kialakulása. Ezen a stádiumon egyszerűen átugrik a fejlődő állat és minden metamorfózis nélkül a felnőtt állat kicsinyített mása jön létre, így több értelme van a fontos sejtkomponenseket oda koncentrálni, ahol igazán kellenek majd. (Vagyis azokba a sejtekbe, amelyekből a kifejlett egyed szövetei jönnek majd létre.)
Ha Tubifex-ről esik szó, akkor jó eséllyel mindenkinek csak az izgő-mozgó, gorgófő-szerű halkaja csomó ugrik be és legkevésbé sem az, hogy ez a féregfaj is a fejlődésbiológia egyik modellállata.
Pedig ez a Kevéssörtéjű férgek csoportjába tartozó állat már hosszú évtizedek óta kitartó figyelmet kapott pár labor részéről, így fejlődéséről is viszonylag sokat tudunk. Standard spirális osztódással indul a történet (akárcsak a pár napja megénekelet csigák esetében), de például a közeli rokon Többsertéjű férgektől eltérően nagyon hamar feltűnik, hogy az első osztódások igencsak aszimmetrikusak, és két igen különböző méretű sejtet hoznak létre. Általában elmondható, hogy a nagyobb sejtből a kifejlett (felnőtt) állat szervez fognak kialakulni, míg a kisebb sejteknek csak kevésbé jól definiálható, obskurus szerep jut. Mindez azért van így, mert az említett Többsertéjűekkel ellentétben itt nem történik meg jellegzetes alakú lárvák kialakulása. Ezen a stádiumon egyszerűen átugrik a fejlődő állat és minden metamorfózis nélkül a felnőtt állat kicsinyített mása jön létre, így több értelme van a fontos sejtkomponenseket oda koncentrálni, ahol igazán kellenek majd. (Vagyis azokba a sejtekbe, amelyekből a kifejlett egyed szövetei jönnek majd létre.)
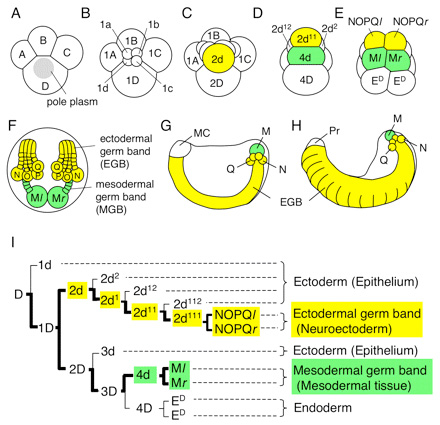 A Tubifex fejlődése, mint az Ősszájúaké általában, igencsak determinisztikus. Ez azt jelenti, hogy a sejtsorsok egyáltalán nem plasztikusak, hanem már az első osztódás után pontosan lehet tudni, hogy a jobboldali és a baloldali sejtből milyen sejtvonalak jönnek létre. (Ez pl. gerinceseknél egyáltalán nem így van, a korai embrió sejtjei teljesen egyenértékűek, minden szempontból.) Olyannyira így van ez, hogy gyakorlatilag számok és betűk kombinációjával pontosan ábrázolhatók a sejtvonalak, leírható gyakorlatilag a teljes embrionális fejlődés, lásd a mellékelt ábra (I) panelje.
A Tubifex fejlődése, mint az Ősszájúaké általában, igencsak determinisztikus. Ez azt jelenti, hogy a sejtsorsok egyáltalán nem plasztikusak, hanem már az első osztódás után pontosan lehet tudni, hogy a jobboldali és a baloldali sejtből milyen sejtvonalak jönnek létre. (Ez pl. gerinceseknél egyáltalán nem így van, a korai embrió sejtjei teljesen egyenértékűek, minden szempontból.) Olyannyira így van ez, hogy gyakorlatilag számok és betűk kombinációjával pontosan ábrázolhatók a sejtvonalak, leírható gyakorlatilag a teljes embrionális fejlődés, lásd a mellékelt ábra (I) panelje.
A Tubifex fejlődése során az első két osztódás négy különböző méretű sejtet (ún. kvadránst) hoz létre: A, B, C, D. Ezekből a D a legnagyobb és legfontosabb, hiszen gyakorlatilag ennek az utódsejtjeiből épül majd fel az állat. A D kvadráns (aszimmetrikus) osztódásai hozzák létre az egymás mellett elhelyezkedő 2d és 4d sejteket, amelyekről tudjuk, hogy a mesoderma és az idegrendszert kialakító neuroectoderma kialakításáért felelnek. Ezekről a sejtekről korábban sejtették, hogy az említett folyamatokon túl kulcsszerepet játszhatnak az állat teljes testtengelyének kialakításában, de kísérletes bizonyíték még nem volt minderre.
Most azonban lett: sejtátültetések segítségével sikerült igazolni, hogy az említett két sejt (de csak együtt, külön-külön nem) valóban elengedhetetlen, hogy a Tubifexeknek feje és farka legyen.
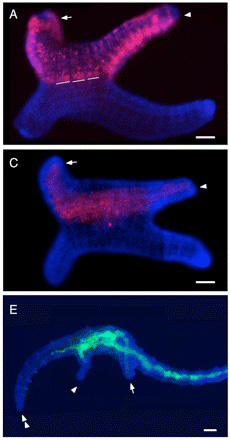
Első körben a két sejtet elpusztították és ekkor csak egy amorf kis gömb jött létre az embrionális fejlődés során, nem egy felismerhető kis féreg (lásd A vs. B, alább). Ha azonban a két sejt elpusztítása után egy másik embrió 2d és 4d sejtjét a helyükre ültették, akkor ismét egy normális testtengelyű egyed jött létre (C).
![]() A legizgalmasabb kísérlet azonban csak ezután jött: nem a 2d és 4d sejteket lőtték ki egy fejlődő embrióban, hanem a túlsó oldalon levő sejtek egyikét (ennek önmagában nincs túl sok következménye, a féreg ugyanúgy cseperedik) és ennek a helyére ültették egy másik embrió 2d és 4d sejtjeit. Az így létrejövő embriónak tehát két-két 2d és 4d sejtje van ellenkező pólusokon. Az eredmény (D) drámai, X és Y alakú egyedek alakultak ki az esetek több mint harmadában, olyan állatok, amelyeknek két farkuk és gyakran két fejük is volt.
A legizgalmasabb kísérlet azonban csak ezután jött: nem a 2d és 4d sejteket lőtték ki egy fejlődő embrióban, hanem a túlsó oldalon levő sejtek egyikét (ennek önmagában nincs túl sok következménye, a féreg ugyanúgy cseperedik) és ennek a helyére ültették egy másik embrió 2d és 4d sejtjeit. Az így létrejövő embriónak tehát két-két 2d és 4d sejtje van ellenkező pólusokon. Az eredmény (D) drámai, X és Y alakú egyedek alakultak ki az esetek több mint harmadában, olyan állatok, amelyeknek két farkuk és gyakran két fejük is volt.
 Akinek volt szerencséje akár csak alapfokú fejlődésbiológiai előadásokhoz, annak erről természetesen Hans Spemann, Hilde Mangold és klasszikus kísérletük ugrik be, ahol gőte-embriókban igazolták, hogy a testtengely indukciója csak néhány mesodermális sejten múlik. Ezek átültetése elég volt ahhoz, hogy a környező sejteket a gazda embrióban arra ösztönözzék, hogy új testtengelyt (izmokkal, idegrendszerrel, belekkel, erekkel) alakítsanak ki.
Akinek volt szerencséje akár csak alapfokú fejlődésbiológiai előadásokhoz, annak erről természetesen Hans Spemann, Hilde Mangold és klasszikus kísérletük ugrik be, ahol gőte-embriókban igazolták, hogy a testtengely indukciója csak néhány mesodermális sejten múlik. Ezek átültetése elég volt ahhoz, hogy a környező sejteket a gazda embrióban arra ösztönözzék, hogy új testtengelyt (izmokkal, idegrendszerrel, belekkel, erekkel) alakítsanak ki.
Tubifex-ben azért nem ennyire drámai a helyzet, de analóg. Sejtsors térképezéssel igazolták, hogy a másodlagos tengely esetében az idegrendszer az átültetett 2d sejtből jött létre a mesoderma pedig kizárólag az átültetett 4d-ből. (Az átültetett sejteket piros festékkel jelölték, és látszik, hogy a másodlagos tengely releváns szövetei mind megfestődtek, mind a 2d (A), mind a 4d (C) jelölése esetében.)
Az egyetlen csíralemez, amit valóban a gazdaszervezet sejtjei alakítanak ki, az az endoderma. Ebben az esetben a gazdaembrió enodermát létrehozó sejtjeit jelölték zöld festékkel, és jól észrevehető, hogy mind a két kialakuló tengelyben a cső alakú emésztőrendszert (ami endodermális eredetű) zöld sejtek hozzák létre.
A felszíni hasonlóságok mellett túl sok különbség van itt ahhoz, hogy azt komolyan fel lehessen vetni, hogy a gerincesek és a gyűrűsférgek embrionális testtengelyeit kialakító „szervezőközpontok” közös eredetűek. De mindenesetre érdekes, hogy analóg mechanizmusok működnek a testszervezés során, ennyire távoli rokonságban levő szervezetekben is.
Nakamoto A, Nagy LM, Shimizu T. (2011) Secondary embryonic axis formation by transplantation of D quadrant micromeres in an oligochaete annelid. Development 138(2): 283-90.