
 Mutációk vannak, és ennek ezer bizonyítékát látjuk, például a gyorsan változó vírustörzsekben, a rezisztenssé váló baktériumokban. Szekvencia szinten is be tudjuk azonosítani őket, akár egymást követő generációkban, és különböző ötletes kísérletekkel meg tudjuk határozni az egyes élőlények ún. mutációs rátáját is, vagyis azt a gyakoriságot, hogy egy-egy mutáció rögzüljön.
Mutációk vannak, és ennek ezer bizonyítékát látjuk, például a gyorsan változó vírustörzsekben, a rezisztenssé váló baktériumokban. Szekvencia szinten is be tudjuk azonosítani őket, akár egymást követő generációkban, és különböző ötletes kísérletekkel meg tudjuk határozni az egyes élőlények ún. mutációs rátáját is, vagyis azt a gyakoriságot, hogy egy-egy mutáció rögzüljön.
Bár kétségtelen, hogy a mutációk léte és örkölhetősége az evolúció egyik hajtóereje, az is biztos, hogy a legtöbb mutáció gondot okoz a gazdájának, adott esetben letális is lehet, ezért a legtöbb élőlény igyekszik minimalizálni a mutációk számát, különböző molekuláris mechanizmusok révén. A legtöbb DNS másolást végző enzim, azaz polimeráz, beépített hibajavító mecahnizmusokkal rendelkezik, s ha valami átcsúszna ezen a szitán még mindig akadnak másodlagos javító rendszerek.
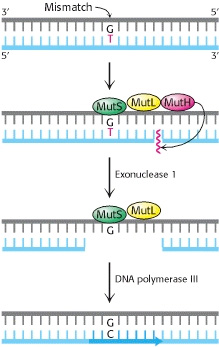
Jobb oldalon az Escherichia coli baktérium hibajavító rendszere látható (a kép innen származik), amelynek logikája a következő: ha a DNS másolásba hiba csúszott, a rosszul párosodó bázispárhoz a MutS nevű fehérje fog kapcsolódni, ami ekkor egy másik fehérjét, a MutL-t is képes lesz megkötni. Utóbbi pedig a MutH-t vonza a probléma helyszínére. Ez egy szakaszon elemészti az újonnan szintetizált, mutációt hordozó DNS-t és a létrejövő lyukat a polimeráz betömi.
A rendszer viszonylag jól működik, csak akkor csúszik hiba a dologba, ha nem elég gyors a javítás: a MutH annak alapján tudja eldönteni, hogy a kettős spirál melyik tagja új, hogy a metilációs mintázatokat figyeli. Az új szál nem metilált, viszont ez csak ideig óráig marad így, mert egy másik enzim előbb-utóbb ráakasztja a metil-csoportokat a megfelelő pozícióba. Ha eddig a pontig a MutH nem végezte el a feladatát, ezután már nem tudja, mert nem tud különbséget tenni a két szál között, így a mutáció megmarad.
Bár a hibás DNS másolás nyomait nem ördöngösség meglelni és olyan egyedek vizsgálatával, ahol a mutációk száma megszaporodik, be tudjuk azonosítani azokat a fehérjéket is, amelyek a sejten belül próbálnak őrködni a genetikai anyag épsége felett, a mutáció kialakulásának pillanatát szinte lehetetlen elkapni. Ez egyrészt technikai jellegű probléma, hiszen hogyan veszed észre, hogy épp mutáció keletkezett, másrészt millió/milliárd bázispár egyszerre történő megfigyelése szintén nem triviális.
Mutációk vannak, és ennek ezer bizonyítékát látjuk, például a gyorsan változó vírustörzsekben, a rezisztenssé váló baktériumokban. Szekvencia szinten is be tudjuk azonosítani őket, akár egymást követő generációkban, és különböző ötletes kísérletekkel meg tudjuk határozni az egyes élőlények ún. mutációs rátáját is, vagyis azt a gyakoriságot, hogy egy-egy mutáció rögzüljön.
Bár kétségtelen, hogy a mutációk léte és örkölhetősége az evolúció egyik hajtóereje, az is biztos, hogy a legtöbb mutáció gondot okoz a gazdájának, adott esetben letális is lehet, ezért a legtöbb élőlény igyekszik minimalizálni a mutációk számát, különböző molekuláris mechanizmusok révén. A legtöbb DNS másolást végző enzim, azaz polimeráz, beépített hibajavító mecahnizmusokkal rendelkezik, s ha valami átcsúszna ezen a szitán még mindig akadnak másodlagos javító rendszerek.
Jobb oldalon az Escherichia coli baktérium hibajavító rendszere látható (a kép innen származik), amelynek logikája a következő: ha a DNS másolásba hiba csúszott, a rosszul párosodó bázispárhoz a MutS nevű fehérje fog kapcsolódni, ami ekkor egy másik fehérjét, a MutL-t is képes lesz megkötni. Utóbbi pedig a MutH-t vonza a probléma helyszínére. Ez egy szakaszon elemészti az újonnan szintetizált, mutációt hordozó DNS-t és a létrejövő lyukat a polimeráz betömi.
A rendszer viszonylag jól működik, csak akkor csúszik hiba a dologba, ha nem elég gyors a javítás: a MutH annak alapján tudja eldönteni, hogy a kettős spirál melyik tagja új, hogy a metilációs mintázatokat figyeli. Az új szál nem metilált, viszont ez csak ideig óráig marad így, mert egy másik enzim előbb-utóbb ráakasztja a metil-csoportokat a megfelelő pozícióba. Ha eddig a pontig a MutH nem végezte el a feladatát, ezután már nem tudja, mert nem tud különbséget tenni a két szál között, így a mutáció megmarad.
Bár a hibás DNS másolás nyomait nem ördöngösség meglelni és olyan egyedek vizsgálatával, ahol a mutációk száma megszaporodik, be tudjuk azonosítani azokat a fehérjéket is, amelyek a sejten belül próbálnak őrködni a genetikai anyag épsége felett, a mutáció kialakulásának pillanatát szinte lehetetlen elkapni. Ez egyrészt technikai jellegű probléma, hiszen hogyan veszed észre, hogy épp mutáció keletkezett, másrészt millió/milliárd bázispár egyszerre történő megfigyelése szintén nem triviális.
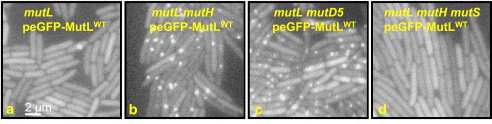 Mégis ezzel kísérletezett most egy csoport, és igen jópofa eredményeket kaptak. A rendszer lényege, hogy egy olyan MutL fehérjét hoztak létre, amihez egy zölden fluoreszkáló fehérje darabot fűztek. Ha ezt az új, rekombináns fehérjét bevitték egy, a mutL génben egyébként mutáns baktérium törzsbe, akkor kb nomrális mutációs rátájú transzgénikus törzs keletkezett. Annyiban különbözött „vad típusú” kollegáitól, hogy időnként egy-egy zöld pont jelent meg egy-egy baktérium kormoszómáján (a), ami idővel eltűnt. Mivel tudjuk, hogy általában több mint egy MutL fehérje köt a pontmutációk környékén, kézenfekvő feltételezni, hogy a zöld pontok pontmutációnak felelnek meg.
Mégis ezzel kísérletezett most egy csoport, és igen jópofa eredményeket kaptak. A rendszer lényege, hogy egy olyan MutL fehérjét hoztak létre, amihez egy zölden fluoreszkáló fehérje darabot fűztek. Ha ezt az új, rekombináns fehérjét bevitték egy, a mutL génben egyébként mutáns baktérium törzsbe, akkor kb nomrális mutációs rátájú transzgénikus törzs keletkezett. Annyiban különbözött „vad típusú” kollegáitól, hogy időnként egy-egy zöld pont jelent meg egy-egy baktérium kormoszómáján (a), ami idővel eltűnt. Mivel tudjuk, hogy általában több mint egy MutL fehérje köt a pontmutációk környékén, kézenfekvő feltételezni, hogy a zöld pontok pontmutációnak felelnek meg.
De a feltételezés még nem bizonyíték, és azt sem tudjuk, vajon mindenféle pontmutáció „bezöldül”, vagy csak azok, amelyek nem javítódnak ki? Ezek eldöntésére két kísérletet végezhetünk. Egyrészt megnézzük a zöld pöttyök („fókuszok”) számát, olyan baktériumokban, ahol a javító mechanizmus valamelyik más eleme elromlik (ez praktikus okokból a mutH kell legyen, mert ha a mutS hiányzik a rendszerből, a fent részletezett okok miatt, a MutL sem tud majd a DNS-hez kötni, lásd (d)), és olyanokban, ahol a polimeráz bépített hibajavítása nem működik (pl. a mutD5 törzs). Mindkét esetben lényegesen megnő a fókuszok száma ((b) és (c)), ami azt jelenti, hogy amit látunk azok a ki nem javított pontmutációk (ha ugyanis a kijavításra kerülők is láthatóak lesznek, akkor a mutH törzsben észlelt fókuszok száma nem különbözhetne nagyon a kontroll csoporttól).
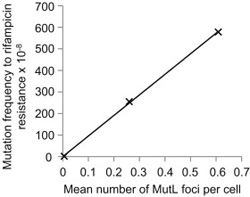 A másik kísérlet pedig annak az összevetése, hogy miképpen korrelál a mutációk száma és a fókuszok száma. Ez pedig lineáris összefüggésnek bizonyult.
A másik kísérlet pedig annak az összevetése, hogy miképpen korrelál a mutációk száma és a fókuszok száma. Ez pedig lineáris összefüggésnek bizonyult.
Azaz, látható fluoreszcens jel csak akkor alakul ki, amikor a létrejövő pontmutációk hosszú ideig megmaradnak, mert a bekövetkező metiláció miatt javításuk nem következhet be (magyarán fixálódnak).
Ugyan a kísérleti rendszer nem teszi lehetővé, hogy lássuk, fizikailag mi is a bekövetkező mutáció, azt élőben követhetjük nyomon, hogy milyen gyakorisággal mutálódik a bakteriális genom.
Elez M, Murray AW, Bi L-J, Zhang X-E, Matic I, Radman M (2010) Seeing Mutations in Living Cells Curr Bio 20: 1432-1437.
Hát ez érdekes volt, tetszett. Most feltennék néhány laikus kérdét:
1. Jól értem, hogy ez a javító mechanizmus csak akkor működik, ha össze nem illő bázisok kerülnek egymással szembe?
2. Ez csak az Escherichia coli-ra jellemző folamat?
3. Ha bekövetkezik a metiláció (bármi is legyen az:)), akkor az ilyen jellegű hibák rögzülenk. Ezek szerint, lehet, hogy a baktérium genomjának bizonyos részei tele vannak öszze nem illő bázisokkal?
4. Az is érdekelne, hogy más jellegű mutációkat milyen mechanizmusok javítanak ki…bár gondolom, kellene ahhoz néhány poszt, hogy ez kifejtsd.
Bocs a sok kérdésért! Az is jó, ha csak belinkelsz valamit, ahol megtalálom a válaszokat. Előre is köszönöm!
1. igen
2. nem, más bacikra is, és a mutS-mutL rendszerhez hasonló fellelhető eukariótákban, csak ott az utolsó lépés megy másként, nem metiláció függően.
3. A metiláció során egy-egy metil (-CH3) csoport kötődik bizonyos bázisokhoz. Ez kb egyenletesen következik be az újonnan megszintetizált szálakon, így ettől a mechanizmustól – amennyire tudom – nem lesznek olyan régiói a baktériumnak, ahol gyakoribbak lennének a rögzült mutációk.
4. Hajaj, ahhoz egy könyvnyi fejezetet kellene írnom. De ha érdekel, akkor itt kezd az olvasgatást:
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=cell&part=A1023
http://www.be-md.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=genomes&part=A8349