
 A gerincesek szemének kifejlődéséről konkrétan könyvet lehet írni, így ha a lényeget egyetlen posztba akarom besűríteni, az akaratlanul is egyszerűsítésekkel és általánosításokkal jár. (Remélem egyik sem megy az érthetőség róvására, de ha igen tessenek majd kérdezni.)
A gerincesek szemének kifejlődéséről konkrétan könyvet lehet írni, így ha a lényeget egyetlen posztba akarom besűríteni, az akaratlanul is egyszerűsítésekkel és általánosításokkal jár. (Remélem egyik sem megy az érthetőség róvására, de ha igen tessenek majd kérdezni.)
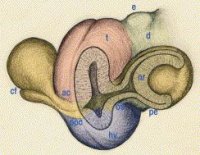
A gerinces állatokra jellemző szemgolyó hasonló képalkotási elvvel működik, mint egy fényképezőgép: egy lencsén keresztül gyűjti be a fénysugarakat és a szem hátsó részén hoz létre egy fordított, kicsinyített képet. Ennyiben hasonlít egyes oktopuszok szemére is, de a hasonlóság csak igen távoli: mind a fejlődés lépéseit, mind a szem egyes sejttípusainak eredetét tekintve jelentős különbségeket találunk. Ez az összehasonlítás önmagában is megér egy ódát (igyekszem egyszer alkalmat is keríteni rá), de most a részletek ismertetése nélkül legyen elég annyi, hogy a különbségek azt bizonyítják, hogy az evolúció különböző módokon is képes volt létrehozni nagyon hasonló működésű, összetett szerveket.
A gerincesek szemfejlődése három nagy fázisra osztható (a felosztás kicsit önkényes, egyszerűbb ezeket követve tárgyalnom a folyamatot – természetesen más felosztások is létezhetnek, a lényeg ugyanaz marad).
Az első fázis a szemmező kialakulása, amely időben átfed azzal a folyamattal, amely során az ektodermális csíralemez egyik része elkülönül, létrehozva az ún. neuroektodermát, amelyből később az idegrendszer jön majd létre. 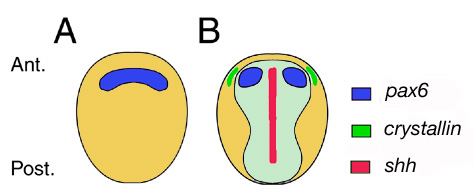 Az egybeesés egyáltalán nem véletlen, hiszen a gerincesek szeme durva egyszerűsítésben (mint azt rögtön látni fogjuk) az előagy kitüremkedése. Ezért, amíg nem kezdenek elkülönülni az idegrendszert alkotó sejtek, addig értelmetlen is a szemről beszélni.
Az egybeesés egyáltalán nem véletlen, hiszen a gerincesek szeme durva egyszerűsítésben (mint azt rögtön látni fogjuk) az előagy kitüremkedése. Ezért, amíg nem kezdenek elkülönülni az idegrendszert alkotó sejtek, addig értelmetlen is a szemről beszélni.
A szemmező a neuroektoderma legelején (Ant.) alakul ki és a legelső gének egyike, ami ezen a területen kifejezésre kerül az a pax6 gén, amely az élővilágban univerzálisan (laposférgektől, a muslicákon át, az emberekig) a szemkialakulásért felel (A). Először a pax6 egy félkör alakú sávban expresszálódik, ez a tartomány azonban hamarosan kettéválik, az embrió középvonalában, az ektodermális réteg alól, a mezodermából érkező jelek hatására (B). Ezen jelek közül legfontosabb a Sonic-hedgehog (Shh) nevű – igen, a genetikusok szeretnek hülye neveket adni 😉 – nevű fehérje, amelynek mutációja, vagy működésének valamilyen külső okból történő gátlása a szemmező szétválásának elmaradásához vezet, ami egyszeműséget, azaz küklopsziát okoz. (Például a cyclopamine nevű molekula egy ilyen Shh működést gátló teratogén anyag és mivel egyes növényekben különösen magas koncentrációban van jelen, az ezeket legelő vemhes tehenek gyakran egyszemű borjaknak adnak életet.)
A fejlődés második fázisa a neurulációt követően következik be. Ez utóbbi az a folyamat, amelynek 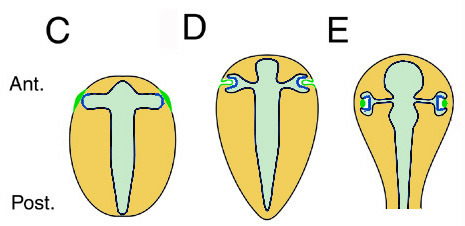 során a neuroektoderma, ami eddig egy lényegében egy sima lemez, csővé – ún. velőcsővé – hajlik össze és besüllyed a leendő kültakaró alá. Ekkor a velőcső legelején, amely az előagyat hozza majd létre, két oldalt kidudorodások jelennek meg, a szemhólyagok. Ezek fokozatosan növekednek, mígnem elérik a kültakaró oldalsó részét (C). Itt a kültakaró (amelynek ezen része már korábban létrehozta a krisztallin géneket kifejező ún. lencse-placodot) és a szemhólyag között bonyolult, és részleteiben máig sem teljesen tisztázott kölcsönhatássorozat indul meg. Ennek az az eredménye, hogy a lencse-placod lefűződik és létrehozza a szemlencsét, a szemhólyag közepe pedig visszatűrődik, így létrejön a szemserleg (D, E). (Ugyanez, keresztmetszeti mikroszkópos képekben itt látható.) Mindeközben a szemhólyag oldalsó sejtjei, amelyekből később a szemserleg visszatűrődő sejtjei lesznek, továbbra is termelik a Pax6-t. Ezek a Pax6+ sejtek hozzák létre később a retinát, a szemserleg őket körülvevő (külső) sejtjei pedig pigmentsejtekké válnak.
során a neuroektoderma, ami eddig egy lényegében egy sima lemez, csővé – ún. velőcsővé – hajlik össze és besüllyed a leendő kültakaró alá. Ekkor a velőcső legelején, amely az előagyat hozza majd létre, két oldalt kidudorodások jelennek meg, a szemhólyagok. Ezek fokozatosan növekednek, mígnem elérik a kültakaró oldalsó részét (C). Itt a kültakaró (amelynek ezen része már korábban létrehozta a krisztallin géneket kifejező ún. lencse-placodot) és a szemhólyag között bonyolult, és részleteiben máig sem teljesen tisztázott kölcsönhatássorozat indul meg. Ennek az az eredménye, hogy a lencse-placod lefűződik és létrehozza a szemlencsét, a szemhólyag közepe pedig visszatűrődik, így létrejön a szemserleg (D, E). (Ugyanez, keresztmetszeti mikroszkópos képekben itt látható.) Mindeközben a szemhólyag oldalsó sejtjei, amelyekből később a szemserleg visszatűrődő sejtjei lesznek, továbbra is termelik a Pax6-t. Ezek a Pax6+ sejtek hozzák létre később a retinát, a szemserleg őket körülvevő (külső) sejtjei pedig pigmentsejtekké válnak.
A harmadik fázis magának a retinának a létrejötte. (Természetesen még ezer egyéb folyamat is zajlik eközben, pl. a szemlencse integrálódása, a szem erezetének kialakulása, szemmozgató izmok megjelenése, stb., de mint említettem, helyhiány okán ezekről most nem lesz szó.) Az előbb említett Pax6+ sejtek osztódásai során jön létre egy jellegzetes, ötrétegű szövet: legbelül (a fénytől legtávolabb eső oldalon) helyezkednek el a fényérzékelést végző fotoreceptorok (csapok és pálcikák), kifele pedig egymás után a horizontális sejtek, bipoláris sejtek, amacrin sejtek és végül a retinális ganglion sejtek (RGC). Utóbbiak hosszú nyúlványai, axonjai aztán szépen kilépnek a szemből az ún. vakfolton keresztül (amelynek neve a fényérzekelő sejtek hiányára utal – ezek fizikailag nem lehetnek ott, az axonok miatt), létrehozva a látóideget, és az agyba továbbítják a csapokból és pálcikákból érkező impulzusokat.
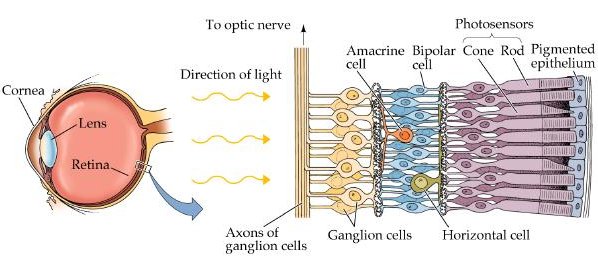
A (gerinces)retinát tehát egy furcsa fordítottság jellemzi, ami egyben azt is jelenti, hogy a fotonoknak több sejtrétegen, illetve axonok kusza sokaságán kell átküzdeniük magukat, mielőtt fényérzetet keltenek. Ezért nevezzük a gerincesek szemét „inverz szemnek”, szemben a korábban már emlegetett oktopuszok szemével, amely egy „everz szem”, vagyis a retina fényhez legközelebbi részén helyezkednek el a fotoreceptorok. (Nem bizonyítottan, de valószínűsíthetően, az „inverz szem” fordítottsága az idegrendszer központosulása miatt alakult így evolúciósan. Ui. a központosulásban kulcsszerepet játszik a neuruláció folyamata, amikor a létrejövő velőcső kialakulása során a korábban a neuroektodermából „kifelé” néző sejtek, a cső belseje fele kezdenek nézni.)
Egy másik érdekesség, amiről még ildomos említést tenni, az a fotoreceptorok működéséhez kapcsolódik. Ellentétben ugyanis számos „mezei” idegsejttel (illetve, a nem-gerincesek szerkezetében picit más fotoreceptoraival), a csapok és pálcikák épp akkor nem továbbítanak jelt, amikor izgalmi állapotba kerülnek. A fotoreceptorok ugyanis sötétben folyamatosan „tüzelnek”, ám amikor erős fény éri őket, akkor hiperpolarizálttá válnak, vagyis rövid időre „elhallgatnak”. Ez picit paradoxnak tűnhet, de az idegrendszer számára nem az a fontos, hogy miképp tesznek érzékszerveink különbséget a fény és sötétség között, hanem csak az, hogy valamiképpen különbséget tegyenek. A többi, azaz a kép értelmezése és a válaszreakció megalkotása, már az agyunk feladata.
Nem terveztek esetleg egy postot a szem evolúciós kialakulásáról is? Különösen, hogy ez meglehetősen kurrens téma volt a kreacionizmus idején: sokáig az volt az elfogadott nézet, hogy a szem annyira bonyolult rendszer, és annyira függ minden egyes része más részektől, hogy nem alakulhatott ki lépésről lépésre, csakis egyszerre, eyg teremtő jóvoltából. A wikipedia-n egyébként egy jó cikk van erről (http://en.wikipedia.org/wiki/Evolution_of_the_eye), de talán ti is hozzá tudtok még tenni valamit.
Remenyeim szerint elobb-utobb lesz (ertsd valszleg nem a kozeljovoben), mert a pl. gerinctelenek szemenek evolucioja eleg jol kovetheto.
Dan-Erik Nilsson volt az, aki csinált egy olan modelt, ami természetes szelekcióval alakítja ki a szemet. Ez az ő honlapja: http://www.biol.lu.se/funkmorf/vision/dan/Dan.html
Itt van egy videó a modelről:
http://www.pbs.org/wgbh/evolution/library/01/1/l_011_01.html
És itt ír róla Dawkins:
http://www.simonyi.ox.ac.uk/dawkins/WorldOfDawkins-archive/Dawkins/Work/Articles/1995-06-16peepers.shtml
Persze ez sem tetszik a kreacionistáknak:
http://www.reviewevolution.com/press/pressRelease_EyeEvolution.php