
A kétoldali szimmetriájú állatok (és a jelek szerint nemcsak) minimum két szimmetria tengely mentén oszthatók fel: az egyik a test hosszán végigfutó anterior-posterior (AP) tengely, a másik pedig a hát-has viszonylatban felfedezhető dorso-ventrális (DV) tengely. Az előbbiről már többször írtam a Hox gének kapcsán, most azonban egy kicsit az utóbbival szöszmötölnék.
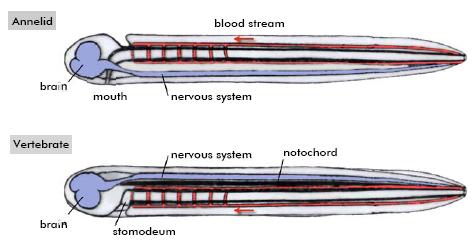 Míg az AP tengely viszonylag világosan és racionálisan definiálható, addig maga a DV tengely definíciója mindig egy kicsit ad hoc-abb jellegű volt: dorsális oldal mindig a föld felőli (ez általában megegyezik a száj pozíciójával is), a ventrális pedig az ég felőli.
Míg az AP tengely viszonylag világosan és racionálisan definiálható, addig maga a DV tengely definíciója mindig egy kicsit ad hoc-abb jellegű volt: dorsális oldal mindig a föld felőli (ez általában megegyezik a száj pozíciójával is), a ventrális pedig az ég felőli.
Egy ilyen definíció alapján, azonban – mint az már a 19. századi zoológusoknak is feltűnt – az ízeltlábúak és a gerincesek között egy érdekes ellentét van: előbbieknek dorsális központosított idegrendszere és ventrális szíve van, míg utóbbiaknak éppen fordítva. Ez a megfigyelés ihlette Étienne Geoffroy St. Hilaire híres "inverziós elméletét", amely szerint a gerincesek őse az evolúció során dorsoventrálisan "megfordult", majd a szája ismét a ventrális oldalra került.
Sokáig úgy tűnt, hogy a molekuláris adatok is egyértelműen alátámasztják a francia zoológus korabeli intuícióját. A DV tengelyt molekulárisan a Bone Morphogenetic Protein (BMP) fehérjék és azokak antagonistái (pl. Chordin, Noggin) közötti kölcsönhatás hozza létre. Az egyik csoport a tengely egyik végén kerül expresszióra, a másik pedig a szemben levő oldalon. Az ízeltlábúaknál a BMP-k dorsálisan vannak jelen, míg anatagonistáik ventrálisan, a gerinceseknél pedig éppen fordított a helyzet, szinte álomszerűen igazolva St. Hilaire-t. Ráadásul ezen két molekulacsoport kölcsönhatása egyértelműen fontos az idegrendszer centralizációjában, így hamarosan szinte tényként volt elkönyvelve, hogy a központosított idegrendszer már a gerincesek és ízeltlábúak közös ősében is fellelhető volt.
A dolgok harmóniájába azonban alaposan bezavart a gerincesek egy távoli rokona, egy félgerinchúros, a Saccoglossus kowalevskii. Ennek a féregre emlékeztető állatnak, ugyanis nincs központosított idegrendszere, csak a bőrében elhelyezkedő idegsejt-hálózat, annak ellenére, hogy a fent említett (molekuláris) DV tengely jelen van. Márpedig ez azt jelenti, hogy vagy valamilyen különös okból "elvesztette" a központosítást (de mivel egyébb vonatkozásokban a DV tengely remekül működik, sokak szerint ez kevéssé valószínű), vagy az csak a félgerinchúros-gerinchúros szétválás után alakult ki az utóbbi vonalban.
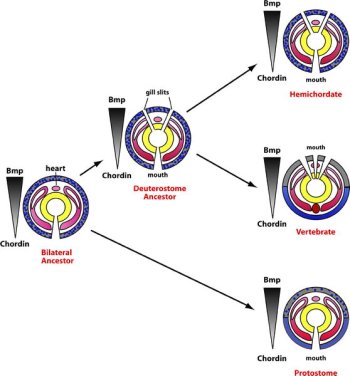 Első pillantásra úgy is tűnhet, hogy ha beigazolódna, hogy az újszájúak (gerinchúrosok, félgerinchúrosok, tüskésbőrűek) és ősszájúak (ízeltlábúak és különböző férek) őse mégsem rendelkezett központosított idegrendszerrel, az tudománytörténeti érdekességgé degradálja az egész "inverziós hipotézist". De ha jobban meggondoljuk ez nem törvényszerű.
Első pillantásra úgy is tűnhet, hogy ha beigazolódna, hogy az újszájúak (gerinchúrosok, félgerinchúrosok, tüskésbőrűek) és ősszájúak (ízeltlábúak és különböző férek) őse mégsem rendelkezett központosított idegrendszerrel, az tudománytörténeti érdekességgé degradálja az egész "inverziós hipotézist". De ha jobban meggondoljuk ez nem törvényszerű.
Ugyanis, bár kétségtelen, hogy kényelmes vele magyarázni az idegrendszer különböző elhelyezkedését, utóbbi nem követelménye az inverziós hipotézisnek. Éppen ezért veti fel Chris Lowe legutóbbi cikkében, hogy lehet, előbb az inverzió következett be és csak ezt követte az idegrendszer centralizációja. S hogy az a két távoli állatcsoportban egyaránt a BMP-Chd tengely mentén következett be, az mindössze azért van, mert a már meglévő DV tengely (amely egyébként a maga módján szabályozza, hogy a félgerinchúrosok bőrében hol milyen típusú idegsejtek keletkezzenek) volt a legkézenfekvőbb módja ezen evolúciós újjítás bevezetésének. (Minthogy az ízeltlábú és gerinces idegrendszer a központosodás kérdésén túl elég sokban különbözik is egymástól, ez egyáltalán nem elképzelhetetlen.)
Bárhogy is van, a legnehezebb kérdés továbbra is nyitott marad: mi miatt történt maga az inverzió …?
Lowe CJ, Terasaki M, Wu M, Freeman RM, Runft L, Kwan K, Haigo S, Aronowicz J, Lander E, Gruber C, Smith M, Kirschner M, Gerhart J. (2006) Dorsoventral patterning in hemichordates: insights into early chordate evolution. PLoS Biol 4(9): e291.
Gerhart J, Lowe C, Kirschner M. (2005) Hemichordates and the origin of chordates. Curr Opin Genet Dev 15(4): 461-467.
Holland ND. (2003) Early central nervous system evolution: an era of skin brains? Nat Rev Neurosci 4(8): 617-627.