
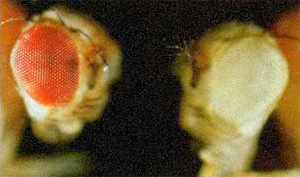
 (Vendégmunkásunk, SexComb újabb posztja.)
(Vendégmunkásunk, SexComb újabb posztja.)
Gyakran esik szó a mutációról,
mint fogalomról, azonban valahogy az az érzésem, hogy sokaknak elég
ködös ezen változások mibenléte, így úgy gondolom, érdekes lehet
egy összefoglaló jellegű írás erről a kérdésről.
Mutációnak nevezzük a sejt
örökítőanyagának megváltozását. Ennyi. Mivel a földi élet
örökítőanyaga a DNS, így a mutációk az adott sejt DNS bázissorrendjének,
azaz szekvenciájának a megváltozását jelentik. Alapvetően három
különböző fajta mutációt különítünk el, ezek a báziscsere,
a deléció és a duplikáció. A báziscsere, más néven pontmutáció,
mint a neve is mutatja, egy nukleotid megváltozása. Teszem azt egy
ACGTGCCAGCTCTTG szakaszból egy pontmutáció során ACGTGCGAGCTCTTG
válik, azaz egy citozin guaninra változott. Deléciónak nevezzük,
amikor az eredeti nukleotidszekvenciához képest hiányoznak bázisok,
mintegy törlődnek. Például a ACGTGCCAGCTCTTG szakaszból
ACGTGAGCTCTTG lesz egy deléció nyomán. Inszerciónak nevezzük
ha az eredeti szekvenciába fölös bázisok épülnek be, például
a ACGTGCCAGCTCTTG szakaszból ACGTGAACCAGCTCTTG lesz egy
inszerció nyomán.
Megkülönböztetünk ezen
kívül kromoszóma-mutációkat, ekkor a DNS szakaszban akkora méretű
változás következik be, amely már a kromoszóma mikroszkópban látható
szerkezetét is megváltoztatja. Mivel a többsejtűek kromoszómái
tulajdonképpen egyetlen meglehetősen hosszú kettősszálú DNS molekulából
állnak, így ezek a mutációk lényegüket tekintve egyedül az érintett
DNS szakasz méretében különböznek a fentebb leírt változásoktól.
(Vendégmunkásunk, SexComb újabb posztja.)
Gyakran esik szó a mutációról,
mint fogalomról, azonban valahogy az az érzésem, hogy sokaknak elég
ködös ezen változások mibenléte, így úgy gondolom, érdekes lehet
egy összefoglaló jellegű írás erről a kérdésről.
Mutációnak nevezzük a sejt
örökítőanyagának megváltozását. Ennyi. Mivel a földi élet
örökítőanyaga a DNS, így a mutációk az adott sejt DNS bázissorrendjének,
azaz szekvenciájának a megváltozását jelentik. Alapvetően három
különböző fajta mutációt különítünk el, ezek a báziscsere,
a deléció és a duplikáció. A báziscsere, más néven pontmutáció,
mint a neve is mutatja, egy nukleotid megváltozása. Teszem azt egy
ACGTGCCAGCTCTTG szakaszból egy pontmutáció során ACGTGCGAGCTCTTG
válik, azaz egy citozin guaninra változott. Deléciónak nevezzük,
amikor az eredeti nukleotidszekvenciához képest hiányoznak bázisok,
mintegy törlődnek. Például a ACGTGCCAGCTCTTG szakaszból
ACGTGAGCTCTTG lesz egy deléció nyomán. Inszerciónak nevezzük
ha az eredeti szekvenciába fölös bázisok épülnek be, például
a ACGTGCCAGCTCTTG szakaszból ACGTGAACCAGCTCTTG lesz egy
inszerció nyomán.
Megkülönböztetünk ezen
kívül kromoszóma-mutációkat, ekkor a DNS szakaszban akkora méretű
változás következik be, amely már a kromoszóma mikroszkópban látható
szerkezetét is megváltoztatja. Mivel a többsejtűek kromoszómái
tulajdonképpen egyetlen meglehetősen hosszú kettősszálú DNS molekulából
állnak, így ezek a mutációk lényegüket tekintve egyedül az érintett
DNS szakasz méretében különböznek a fentebb leírt változásoktól.
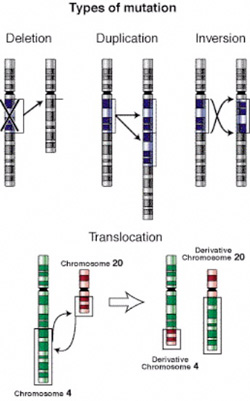 Léteznek
Léteznek
kromoszóma deléciók, ekkor egy kromoszómaszakasz
törlődik. Az érintett szakaszon található
gének természetesen eltűnnek. Léteznek
kromoszóma duplikációk, ekkor egy
kromoszómaszakasz megkétszereződik. Ilyenkor az
érintett szakaszon található gének mind
megkétszereződnek.
Inverziónak
nevezzük, amikor egy kromoszómaszakasz, általában
kromoszómatörések nyomán "megfordul",
azaz maga a kromoszómaszakasz mintegy kivágódik
és fordított állásban forr össze
újra. Transzlokációnak nevezzük azt a
mutációt, amikor egy kromoszómaszakasz letörik
az egyik kromoszómáról és egy másikra
forr oda, az érintett szakasz másik kromoszómára
kerül át. Az inverzió és a transzlokáció
annyiban különleges esetei a kromoszóma mutációknak,
hogy amennyiben a töréspontjaik nem érintenek
egyetlen gént sem, rendszerint nem jelennek meg a
fenotípusban, mivel minden gén megvan, ráadásul
a szükséges példányszámban, így
az adott egyed életképességét nem
befolyásolják, ám ivarsejtképzéskor
komoly gondot okozhatnak. Ezen kívül ismertek egész
kromoszómákat érintő duplikációk
és deléciók is, ilyenkor az adott kromoszóma
a szükségesnél több avagy éppen
kevesebb példányban van jelen a sejtben.
Mi okozza
a mutációkat? Egyrészt maga a DNS lemásolása
sem teljesen hibamentes, a DNS polimeráz enzimek nem
tökéletesek, időnként hibás bázisokat
építenek be, azaz az Adeninnel szemben nem Timint, a
Guaninnal szemben pedig nem Citozint. Ezen kívül néha
mikrodeléciók, illetve kisméretű inszerciók
is történnek a DNS megkettőződésekor a
polimeráz enzim hibájából fakadóan.
Ennek esélye enzimenként változik, például
az E. coli baktérium DNS polimeráz III. enzimje minden
százezredik bázissal szemben nem a neki megfelelő
kiegészítő bázist építi be az
újonnan előállított DNS szálba, azaz
10-5 eséllyel okoz pontmutációt.
Minden sejtben léteznek hibajavító folyamatok,
amelyek ezeket a hibákat felismerik és jó
eséllyel kijavítják, ezek a ténylegesen
bekövetkező mutációk esélyét
10-7 –re csökkentik. Így egyszerűen
a DNS átírása sejtosztódáskor
körülbelül minden tízmilliomodik bázis
mutációját eredményezi. Ez annyit jelent,
hogy mivel az E. coli genom valamivel több, mint négy
megabázis méretű, így hasonló
esélyekkel számolva minden második-harmadik
sejtosztódáskor egy új mutáció
keletkezik magától, minden külső behatás
nélkül. A kólibaktérium 30-40 perces
nemzedékeivel számolva ez azt jelenti, hogy egyetlen
sejtben háromóránként történik
egy új mutáció a polimeráz enzim
hibájából. A sejtmagvasok kromoszómáira
sejtosztódáskor különösen nagy erők
hatnak, ilyenkor fehérjefonalak húzzák a sejt
ellentétes oldalaira őket, időnként
óhatatlanul megsérül egyik-másik, így
kromoszómamutációk is viszonylag gyakran
keletkeznek.
Természetesen
ismertek mutagének is, amelyek nagyságrendekkel növelik
a mutációs gyakoriságot. Például
mindenki hallott már arról, hogy az ibolyántúli
sugárzás képes károsítani a DNSt
és pontmutációkat létrehozni. Nagy
energiájú, ionizáló sugárzások
is képesek károsítani a DNSt, például
a röntgensugárzás gyakran okoz
kromoszómatöréseket, így
kromoszóma-mutációkat. Léteznek mutagén
kémiai anyagok is, például az etil-metán
szulfonát (EMS) báziscsere jellegű mutációkat
okoz. A már korábban tárgyalt P-elemhez hasonló ugráló genetikai elemek is mutagének,
például a P-elem beépülésekor
inszerciós mutációkat okoz, kivágódásakor
pedig gyakran okoz különböző méretű
deléciókat, illetve kromoszómatöréseket.
Ezeken kívül rengeteg mutagén hatás, anyag
létezik, ezeket nem részletezném, elég
sok helyen találni róluk adatokat. Ezeket a hatásokat
semmiképpen sem tudjuk kizárni, hiszen valamennyi
háttérsugárzás folyamatosan éri az
élőlényeket, a táplálékunk
rengeteg különböző szervezet anyagcseretermékeit
tartalmazza, amelyek között több mutagén
ismert, folyamatosan süt minket a nap, ráadásul a
sejt saját energiatermelő folyamatai is veszélyes
szabadgyökök keletkezésével járnak.
Fontos leszögezni, hogy nem különböznek egymástól
a "természetes" és a "mesterséges"
eredetű mutációk. Mindkét esetben
megváltozik a DNS bázissorrendje, ez a változás
az első sejtosztódáskor rögzül, a
maguktól keletkezett mutációk
megkülönböztethetetlenek a mutagén anyagokkal
létrehozott változásoktól, teljesen
egyenértékűek velük, így nincs értelme
"természetes" és "mesterséges"
mutációkról beszélni. (Lásd még itt, itt és itt.)
Nagyon
fontos, hogy egy mutáció kijavítása csak
az első sejtosztódásig lehetséges, amint
bekerült a leánysejtekbe, onnantól azoknak a
sejteknek az a genomja, ezt örökölték, e
szerint működnek. Fontos, hogy milyen sejtben történt
a mutáció. Amennyiben egy ivarsejtben, akkor tovább
adódik az utódnak, ám amennyiben egy testi
sejtben, akkor az ivaros szaporodás során elveszik,
hiszen csak az eredeti mutációs eseményt
elszenvedő sejt leánysejtjei öröklik, az
ivarsejtek nem hordozzák, így nem is adható át
az utódoknak. Az ivarsejtekben megjelenő mutációkat
csíravonal-mutációknak nevezzük, míg
a testi sejtekben történt mutációs
eseményeket szomatikus mutációknak. Amennyiben
egy ivarsejtben történik meg a mutáció, a
belőle kifejlődő utód szervezete semmilyen
módon sem törekszik a mutáció kijavítására,
az ő számára ez a természetes állapot,
nem érzékeli, hogy ő egy mutáns, mivel
nincs semmilyen emléke sem a mutáció előtti
állapotról, amit mi vad típusnak nevezünk.
Például egy cisztás fibrózisban szenvedő
beteg szervezete semmilyen módon sem képes kijavítani
a betegséget okozó mutációt, az ő
számára ez a természetes állapot, még
akkor is, ha belehal.
Az újonnan
keletkezett mutáció hogyan öröklődik?
Egy diploid szervezet, mint amilyen az ember is, minden
kromoszómájából, így minden
génjéből is két példánnyal
rendelkezik, egy apaival és egy anyaival. Amennyiben a
kérdéses allél akkor is kialakítja az ő
fenotípusát, ha csak az egyik kromoszóma
hordozza a kérdéses allélt, a másik a gén
vad típusú változatát tartalmazza, akkor
az adott allél domináns. Azaz ha heterozigóta
állapotban is megjelenik a fenotípusban, a kérdéses
allél domináns. Amennyiben kizárólag
homozigóta állapotban jelenik meg a fenotípusban,
az adott allél recesszív. Az, hogy egy mutáció
domináns vagy recesszív allélt eredményez,
nem lehet előre megmondani, kizárólag az adott
allél ismeretében nyilatkozhatunk róla. Haploid
szervezetek esetében nincs értelme domináns vagy
recesszív mutációkról beszélni,
mivel ilyenkor minden génből csak egyetlen példányt
hordoz az adott szervezet, az egyébként recesszív
mutációk is megjelennek a fenotípusban. Egyedi,
nemhez kötött öröklődést mutatnak az
emlősök X kromoszómáján található
gének. Mivel az emlősök hímjeiben egy X és
egy Y ivari kromoszóma található, a hímek
az ezen kromoszómán kódolt génekből
csak egyetlen példánnyal rendelkeznek, így
esetükben a recesszív allélek is megnyilvánulnak
a fenotípusban. Ilyen például az X kromoszómához
kötött vérzékenység, vagy hemofília.
Ami a fő
kérdés: mit okoz egy adott mutáció? Ezen
a szinten vizsgálva lehetetlen megmondani. Az egyes DNS
szekvencia változások hatása csak akkor válik
egyértelművé, ha megvizsgáljuk, milyen
hatással vannak az adott génre. "Gén"
alatt a genetikai génfogalmat értem, azaz a fehérjét
kódoló DNS szakaszt és az ennek kifejeződését
irányító szabályozóelemeket. Egy
mutáció hatását legjobban a mutációk
Müller-féle csoportosítása ragadja meg.
Hermann Joseph Müller szerint léteznek funkcióvesztéses
és funkciónyeréses mutációk:
- Amorf
mutáció (Funkcióvesztéses): Egy gén
amorf mutációja azt jelenti, hogy a gén által
kódolt fehérje működése teljesen
megszűnik. Lehetséges, hogy a gén szabályozóelemei
válnak működésképtelenné, így
egyáltalán nem is termelődik a fehérjéből,
vagy ha a fehérjét kódoló génszakasz
hibás, megtermelődik ugyan a fehérje, de a
térszerkezete annyira más, mint a vad típusúé,
hogy képtelen ellátni eredeti feladatát. Az
amorf mutációkat szokás még null
mutációknak is nevezni. Amennyiben a fehérje
eredeti működésének ellátásához
elégséges a vad típusban termelődő
mennyiség fele is, az adott amorf mutáció
recesszíven öröklődik, amennyiben a vad
mennyiség fele már kevés a fehérje
eredeti működésének ellátásához,
az adott amorf mutáció dominánsan öröklődik. - Hipomorf
mutáció (Funkcióvesztéses): Egy gén
hipomorf mutációja azt jelenti, hogy a gén által
kódolt fehérje eredeti működése
csökken, de nem szűnik meg teljesen. Ez úgy
lehetséges, hogy a kérdéses fehérjéből
kevesebb termelődik meg, vagy a fehérje szerkezete torzul
és kevésbé működik, mint a vad típusú.
Egy hipomorf mutáció lehet domináns és
recesszív is, az amorfnál leírtak szerint. - Hipermorf
(Funkciónyeréses): Egy gén hipermorf mutációja
annyit jelent, hogy az általa kódolt fehérje
működése megnő a mutáció
hatására, vagy az adott fehérje mennyisége
növekszik meg, vagy a fehérje aktivitása
emelkedik. Egy hipermorf mutáció lehet domináns
és recesszív is. - Antimorf
(Funkciónyeréses): Egy antimorf mutáció
olyan fehérjeterméket eredményez, amely nem
csak, hogy nem képes ellátni eredeti működését,
hanem a vad típusú fehérje működését
is gátolja. Az antimorf mutációk általában
érintik a gén fehérjét kódoló
szakaszát is. Az antimorf mutációk dominánsak.
Gyakran domináns negatívnak is nevezik az antimorf
mutációkat. - Neomorf
(Funkciónyeréses): A neomorf mutációk
esetében az érintett gén fehérjeterméke
valami teljesen más működést mutat, mint a
vad típusú fehérje. Megváltozhat ilyenkor
a fehérje működése, vagy olyan sejtekben is
kifejeződik, amelyekben a vad típusú fehérje
nem. A neomorf mutációk rendszerint dominánsak.
A mutációk
Müller-féle csoportosítása közelebb
visz ugyan az adott mutáció hatásának a
megértéséhez, ám tulajdonképpen
minden egyes mutáció más és más,
nem nagyon mondhatunk róluk általánosságokat.
Éppen ezért szeretném bemutatni egy gén
néhány különböző alléljét.
A kiválasztott gén a Drosophila melanogaster,
azaz az ecetmuslica white génje. A gén a muslica
ivari kromoszómáján helyezkedik el, azaz az X
kromoszómán, így nemhez kötött
öröklődést mutat. Az általa kódolt
fehérje a barna és a vörös szemfesték
termelésévben és a szemsejtekbe juttatásában
játszik szerepet, nem mellesleg a gén w1
allélja volt az első Drosophila melanogasterben
leírt mutáció, még Thomas Hunt Morgan
azonosította 1910 –ben. A white gén számos
allélját azonosították az eltelt száz
év alatt, ezekből szemezgetnék néhányat.
- w1 : Az első azonosított mutáció. Null allél,
azaz a fehérje működése teljesen kiesik, a
Müller-féle felosztás szerint amorf mutáció,
recesszíven öröklődik. A homozigóta
nőstények szeme teljesen fehér. Emberi beavatkozás
nélkül keletkezett, azaz spontán mutáció.
Egy Doc nevű ugráló genetikai elem ugrott a white
génbe, ez tette tönkre teljesen. - wa:
A mint apricot, azaz barack. Ez a white gén egy recesszíven
öröklődő allélje, a homozigóta
legyek szeme halványsárgás, barackszínű,
innen a neve is. Ez a mutáció is emberi beavatkozás
nélkül történt, a white génbe egy
copia ugráló genetikai elem ugrott be, ez csökkentette
le a működését. Az allél nem teljesen
működésképtelen, így a Müller-féle
felosztás szerint hipomorf mutáció, mérések
szerint a vad fehérje aktivitásának csak három
százalékával rendelkezik, ám ennyi is
elég ahhoz, hogy látható mennyiségű
festék jusson az állat szemeibe. - wch:
Ch mint cherry, azaz cseresznye, a homozigóta legyek szeme
rózsaszínes. Ez a mutáció a w1
allélből alakult ki, amikor egy újabb,
pogo típusú ugráló genetikai elem ugrott
be a w1 allélbe. A Müller-féle
felosztás szerint hipomorf allél, recesszíven
öröklődik. Spontán mutáció,
minden emberi beavatkozás nélkül alakult ki. - wBwx:
Bwx mint Brownex, a homozigóta egyedek szeme ugyanis barna.
Heterozigóta formában is megjelenik a fenotípusban,
akinek már nagyon ráállt a szeme, az a
heterozigóta állatokat is felismeri, ám a
homozigóta egyedekben egyértelmű a fenotípusa,
így szemidominánsan öröklődik. A
Müller-féle felosztás szerint hipomorf mutáció.
Spontán alakult ki, emberi beavatkozás nélkül,
a gén fehérjekódoló szakaszában
történt egy rövid deléció. - wDZL:
DZL mint Dominant Zeste-like. Ez a mutáció dominánsan
öröklődik, a heterozigóta hímek szeme
barna, a nőstényeké sárga. Spontán
alakult ki, emberi beavatkozás nélkül. Müller-féle
besorolása antimorf. Úgy keletkezett, hogy két,
egymáshoz közel ülő FB ugráló
genetikai elem ugrott egyszerre a white génbe, így
azonban az ugráskor magukkal hozták az eredeti
beépülési helyük közt lévő
szakaszt a muslica genomból, amely így az eredeti white
génnel együtt íródik át, egy a vad
típusúnál jóval nagyobb méretű
fehérjét kódol. - In(1)wm4h:
Ez az allél egy spontán, emberi beavatkozás
nélkül létrejött kromoszómamutáció
nyomán keletkezett. A Müller-féle besorolása
leginkább hipomorf mutáció, recesszíven
öröklődik. A kromoszómákon a centromer
az a hely, ahová a sejtosztódáskor
összeszerelődő húzófonalak kapcsolódnak.
Éppen ezért a centromer közelében a DNS
szorosan becsomagolt, tömör heterokromatin formában
található, ez valószínűleg azért
szükséges, hogy megfelelően szilárd legyen a
kromoszóma, kibírja a sejtosztódáskor
ébredő erőket. Ennek a heterokromatin régiónak
a kiterjedése egy egyeden belül is változhat kis
mértékben, a pontos határai egyes sejtekben
különböző helyeken találhatóak. Ez
a mutáció egy inverzió, azaz az X kromoszóma
egy szakasza letört a kromoszómáról és
fordított állásban forrt vissza. Ez az
átrendeződés a white gént a
centromer közelébe helyezte át, a heterokromatikus
régió határára, így egyes
sejtekben, ahol a heterokromatikus régió kicsit kisebb
a white gén kifejeződik ezek a szemsejtek pirosak
lesznek, más sejtekben, ahol a heterokromatin kiterjedése
nagyobb, nem fejeződik ki, ezek a szemsejtek fehérek
lesznek. Mindez azt eredményezi, hogy a légy szeme
foltos, azaz fehér alapon apró, piros foltok láthatóak.
Ebben az esetben a white gén ép, ismereteink
szerint a fehérjekódoló szakasz és a
szabályozórégió sem sérült,
egyszerűen a genomon belüli helye változott meg, ez
okozza ezt a fenotípust. - wbl:
Spontán kialakult, recesszív, hipomorf allél. A
wa allélből alakult ki egy újabb
mutációs esemény nyomán, egy blood ugráló
genetikai elem ugrott a white génbe, ez okozza a
fenotípust. Ami miatt érdekes: tizenkilenc
Celsius-fokon tartva az állatokat a szemük színe
sötétvörös, míg harminc Celsius –fokon
tartva az állatokat, a szemük színe barnássárga
lesz. Az ehhez hasonló mutációkat csúnya
szóval kondicionális (feltételes) mutációknak
nevezzük, mivel az általuk kialakított fenotípus
egy külső körülménytől függ. Ez
esetben ez egy hőmérséklet-érzékeny
mutáció, a külső hőmérséklet
változtatásával a fenotípus is
megváltozik.
Nem is
sorolnám tovább, a white génnek több
mint ezer allélját azonosították az évek
során a fenti hivatkozásokon mindenki kedvére
csemegézhet. Ez az ezernél több mutáció
nyilvánvalóan nem az összes létrejött
mutációt jelenti, csak azokat, amelyeket valaki
észrevett, törzsbe állított, és
valahol leközölt. Amit mindezzel meg szerettem volna
világítani: Az egyes mutációk ugyanabból
a génből kiindulva számos különböző
fenotípust képesek kialakítani. Sokat hallani,
hogy a mutációk csak jól behatárolt
változásokat képesek létrehozni. Hol
vannak ezek a határok? Például a white
gén esetében mik a változásnak azok a
határai, amelyek már nem léphetőek át?
Ebből az egy génből, amely által kódolt
fehérje egy jól behatárolt folyamatot katalizál,
különböző mutációs események
rendkívül eltérő fenotípust adó
mutánsokat alakítottak ki. Gyakorlatilag minden olyan
génre, amelyet alaposan vizsgáltak számos
különböző mutáns allélt
azonosítottak, soha semmilyen bizonyítékot sem
találtak bármilyen határra, amelyet a mutációk
által létrehozott változékonyság
ne tudna átlépni. Megfigyelhető, hogy az egyes
mutáns allélek sem védettek a további
változásoktól, olyan allélek is ismertek,
amelyek egy törzsbe állított mutáns allél
újbóli mutációjával keletkeztek,
ezt csak azért szögezném le, hogy elejét
vegyem az olyanfajta elmélkedéseknek, miszerint ezek a
mutációk mindig is megvoltak a természetben,
csak a kutatók homozigóta állapotba hozták
a recesszív alléleket. Kijelenthetjük, hogy az
élőlények genomja változik. Mutációk
folyamatosan történnek, új allélek
folyamatosan alakulnak ki.
A második
központi kérdés, hogy egy mutáció
hasznos avagy káros? Erre a kérdésre a
legnehezebb válaszolni, ugyanis egy mutáció
előnyei és hátrányai csak a környezet
függvényében értelmezhetőek. Nézzünk
néhány emberi példát: A laktóz,
más néven tejcukor egy összetett cukor, ebben az
alakjában csak akkor hasznosul, hogyha lebomlik egy
galaktóztra és egy glükózra, amelyek már
a bélből felszívódnak. A laktóz nagy
mennyiségben található az emlősök
tejében, éppen ezért az emlősök
kicsinyei egy ideig termelnek egy laktáz nevű enzimet,
amely képes egyszerű cukrokká bontani a tejcukrot.
Az emlősök általában csak addig termelnek
laktáz enzimet, amíg szopnak, az elválasztás
ideje után megszűnik ezen enzim termelése is. Az
emberi faj nagy része, a földön élő
hétmilliárd ember háromnegyede esetében
így is történik, négy-hat éves
korukban megszűnik a laktáz enzim termelése,
ezáltal a tejcukor emésztésének képessége
is. Ez után ha az ember tejet fogyaszt, az emésztetlen
tejcukor hasmenést, fokozott gázképződést,
bélgörcsöket okoz, szóval roppant
kellemetlen. Ezt az állapotot laktózérzékenységnek
nevezik, ami öröklött jelleg, az érintett
génnek Európában két allélja
ismert, a domináns allél a laktáz enzim egész
életen át megmaradó termelését,
ezáltal a tejivás képességét
okozza, a recesszív allélre homozigótákban
csak négy-hat éves korukig termelődik laktáz
enzim, utána megszűnik ennek a fehérjének a
termelése, ezáltal a tej fogyasztásának a
képessége is. A két allél közül
a recesszív a vad típusú, amely nem teszi
lehetővé a tejivást, nyilván ez az ősi
változat. Jelenleg is elterjedtebb, mint a tejivást
engedő változat, azon kívül a többi
emlősállatnál jóformán ismeretlen ez
a jelleg. Az alléleloszlás meglehetősen
egyenetlen, például Hollandia lakosságának
csak 1% -a laktózérzékeny, Kína
lakosságának 93% -a ebben a boldog állapotban
leledzik, míg a bennszülött amerikaiak 100% -a
képtelen tejet emészteni. A magyar lakosság 38%
-a laktózérzékeny, mi a középmezőnybe
tartozunk. A laktáz enzim egész életen át
termelése nyilván egy mutáció eredménye.
A korai emberi életkörülmények között
ez a mutáció semleges volt. A vadász-gyűjtögető
emberek, mint ahogy a mai természeti népek is,
semmilyen formában sem fogyasztanak tejet, az ugyanis mint
táplálék csak az állattenyésztés
kezdete után jelenik meg. Az állattenyésztés
kialakulása előtt ez a mutáció semleges, ám
az állattenyésztés, egy külső, nem
biológiai körülmény kialakulása után
ez a mutáció egyértelműen előnyös,
hiszen onnantól kezdve a pásztornak nem kell levágnia
a tehenet, hogy ehessen belőle, elég, ha megfeji. Viszont
onnantól, hogy az ember megkezdte a pásztorkodást,
hamarosan kitalált olyan módszereket is, amelyekkel
sikerült felülemelkednie ezen az örökletes
hátrányon. A joghurt, a kefír, a kemény
sajt ugyanazon az elven alapszik: A mikróbák először
a legkönnyebben felhasználható tápanyagokat
fogyasztják el a környezetükből, a cukrokat, a
tej esetében ez a tejcukor, így ha baktériumokkal
és/vagy gombákkal erjesztik a tejet, azok jelentősen
lecsökkentik a tejcukor-tartalmát, olyan mértékig,
amikor már a tejcukor-érzékeny ember is
biztonsággal megihatja. Akkor most a laktáz enzim egész
életen át termelése előnyös,
hátrányos, vagy semleges mutáció? A
válasz: a körülményektől függ. Az
állattenyésztést nem ismerő környezetben
semleges, tejet fogyasztó környezetben előnyös,
inkább sajtot/kefírt/joghurtot fogyasztó
környezetben semleges. (Erről a mutációról
már írtunk korábban itt és itt.)
Vizsgáljunk
meg egy másik mutációt, a sarlósejtes
vérszegénységet. (Szintén esett már róla szó.
) Ez a mutáció a hemoglobin molekula β-globin
láncát kódoló génben következik
be. Maga a mutáció recesszív, egyetlen bázis
cseréje okozza, a heterozigóták vére
működőképes, ám a homozigóták
vörösvértestjei torzult alakot vesznek fel és
könnyen széttörnek, így a kisebb átmérőjű
erekben gyakran károsodnak, időnként
összetorlódnak, elzárják az apróbb
ereket, ezzel komoly károkat okozva. A mai orvosi ellátás
mellett ez a betegség nem feltétlenül halálos,
ám a betegek időről-időre orvosi segítségre
szorulnak és így is csökken valamelyest a várható
élettartamuk. A mai orvoslás előtt sokkal
komolyabb volt ez az állapot, sokan belehaltak az erek
elzáródásába és az ezzel járó
lázrohamokba. Ez a mutáció ezek alapján
egyértelműen hátrányos, ha meg is jelenik
egy mutációs esemény nyomán, ki kellene
szelektálódnia a populációból,
hiszen csökkenti az egyed életképességét.
Ennek megfelelően Európában rendkívül
ritka, negatív szelekció hat rá. No most
felfigyeltek rá, hogy Afrikában viszont rendkívül
gyakori a sarlósejtes vérszegénység.
Miért különbözik ennyire az eloszlása a
két népességben? Miért szaporodott fel
ennyire ez a káros mutáció Afrikában? A
válasz abban keresendő, hogy ezeken a területeken
fertőz a malária, amely élősködő
életciklusának egy részét a
vörösvértestek belsejében tölti, így
bújik el az emberi immunrendszer elől. A sarlósejtes
vérszegénységre heterozigóta emberek
vörösvértestjei, bár az eredeti feladatukat
képesek ellátni, szétrepednek, ha a malária
élősködője őket fertőzi, így
az élősködő képtelen elbújni az
emberi immunrendszer elől. Ezek a heterozigóta (Aa)
emberek ellenállóak a maláriával szemben,
ám egyébként egészségesek. A malária ma is
gyakori halálok Afrikában, így az ennek
ellenálló egyedek hatalmas előnybe kerültek. Ebben az esetben, ha egy
szülőpár mindkét tagja hordozó, ők
ellenállóak a maláriával szemben, a
gyermekeik negyede vad típusú, fele hordozó,
azaz szintén ellenálló, negyede pedig
sarlósejtes vérszegénységben szenved.
Ebben a környezetben a malária elleni védettség
előnye messze felülmúlja a sarlósejtes
vérszegénységben szenvedő utódok
születéséből származó hátrányt,
így ebben a környezetben pozitív szelekció
hat erre az allélre, így ez a mutáció a
kialakulása után elterjed a populációban.
Ebben az esetben a sarlósejtes vérszegénységet
okozó mutáció maláriamentes területeken
hátrányos és ritka, maláriával
fertőzött területeken előnyös és
gyakori. Akkor ez most előnyös, vagy hátrányos
mutáció?
Egy másik
emberi példa a CCR5 gén és az ez által
kódolt CCR5 fehérje. Ez egy sejtfelszíni
receptorfehérje, amely immunsejtek felszínén
található. Valószínűleg a
gyulladásos immunválaszban játszik szerepet, ám
ez még nem teljesen tisztázott. A CCR5 génnek
létezik egy mutáns allélje, a CCR5-D32, amely
egy rövid, 32 bázispár méretű
delécióval keletkezett, amely tönkreteszi a gén
által kódolt fehérjét. A homozigóta
és a heterozigóta emberek is egészségesek,
így valószínűleg a szervezet valahogyan
képes pótolni a kieső működéseket.
Ez a mutáns allél meglepően gyakori Európában,
a kaukázusi rassz képviselőiben, magyarán
szólva a fehér emberekben. Ami miatt érdekes: Ez
a CCR5-D32 mutáció véd a HIV fertőzés
ellen, a heterozigóták kisebb eséllyel
fertőződnek meg a HIV vírussal, míg a
homozigóták csak kivételes esetekben kapják
el a vírust, gyakorlatilag ellenállónak
tekinthetőek. Ellenben úgy tűnik, hogy a homozigóták
érzékenyebbek a Nyugat-Nílusi Vírus
fertőzésre. Most akkor a CCR5-D32 mutáció
hasznos, vagy káros? HIV fertőzött környezetben
hasznos, Nyugat-Nílusi Vírus fertőzött
környezetben káros. Akkor ez most egy előnyös,
vagy hátrányos mutáció?
Valós
mutációk vizsgálatával megvilágítható,
hogy az az állítás, miszerint hasznos mutációk
nem léteznek többek között azért sem
igaz, mert egy mutáció hasznossága vagy
károssága csak a környezet függvényében
értelmezhető. Egy mutáció, ami egy adott
időben, adott körülmények között
hasznos, egyáltalán nem biztos, hogy egy másik
élőhelyen, más körülmények között
is előnyös. Érdemes elgondolkodni azon, hogy a
Földön hol állandóak a körülmények
tartósan? Alig tízezer éve ért véget
az utolsó jégkorszak, és máris nyakunkon
a globális felmelegedés. A jégkorszak is
gyökeresen átalakította a Föld éghajlatát,
minden élőlénynek alkalmazkodnia kellett a
lehűlésekhez és a felmelegedésekhez is és
ezzel együtt minden más éghajlati változáshoz
is. A Földön nincsenek állandó élőhelyek,
az egyes élőlények környezete folyton
változik, így az egyes különböző
allélek hasznossága vagy károssága
folyton átértékelődik egy adott populáción
belül is. Ezen kívül az éghajlat mellett
számos tényező gyakorolhat szelekciós
nyomást egy adott populációra. Hogy emberi
példát mondjak: Amerika felfedezése után
nem sokkal nagyon erős, addig ismeretlen szelekciós
nyomás alá kerültek az ott élő népek,
ugyanis a fehérek által behurcolt járványok
szinte megtizedelték az őslakókat, akik ezek ellen
gyakorlatilag teljesen védtelennek bizonyultak. Itt például
alig néhány év alatt gyökeresen
megváltozott ezen népek környezete és a
rájuk ható szelekciós nyomás is.
Egy
harmadik fontos kérdés, hogy minden mutáció
megjelenik-e a fenotípusban, vagyis az örökítőanyag
módosulása követhető –e az adott
élőlény megfigyelésével? A válasz
egyszerű: nem. Számos esetben a mutációnak
semmilyen látható vagy mérhető jele sincs.
A genetikai kód degenerált, azaz egy aminosavat több
különböző bázishármas is kódolhat.
Például a GCT, GCC, GCA, GCG bázishármasok
is Alanin aminosavvá fordítódnak le, így
ha egy pontmutáció következtében például
a GCC bázisokból GCA lesz, semmi hatása sem lesz
a fehérje működésére, mivel a
polipeptidlánc nem változik, ugyanaz az aminosav épül
be a kérdéses helyre. Ezen kívül egy-egy
aminosav megváltozása nem feltétlenül teszi
működésképtelenné a fehérjét,
sok olyan pontmutációt, rövidebb deléciót
írtak le, amelyek, bár megváltoztatják a
fehérje aminosavsorrendjét, a működését
nem befolyásolják. A kromoszómamutációk
között tárgyaltuk az inverziókat és a
transzlokációkat. Ilyen kromoszómaátrendeződések
jelenléte nem feltétlenül evidens, gyakran csak alacsonyabb termékenységben nyilvánul meg.
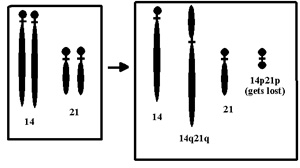 A Down-kórt
A Down-kórt
leggyakrabban a 21. kromoszóma duplikációja
okozza, azaz ezeknek a betegeknek kettő helyett három 21.
kromoszóma található minden egyes sejtjükben.
Azonban létezik egy másik formája is a
Down-kórnak, a Robertson-féle transzlokáció.
Ekkor a 21. kromoszóma hosszú karja letörik és
egy másik kromoszóma valamelyik karjához forr
oda. (Példaként vegyük a 14. kromoszómát)
E mutánsok általában semmilyen fenotípust
sem mutatnak, hiszen minden génjükből egészen
pontosan két példánnyal rendelkeznek. Azonban
ivarsejtképzéskor a kromoszómák
szétválnak. Ha az utód az ép 21. és
a transzlokációval megnövekedett 14. kromoszómát
örökli a szülőtől, akkor a 21. kromoszóma
transzlokáció által érintett karja három
példányban található meg a sejtjeiben,
így kialakul nála a Down-kór. Amennyiben a
"hosszú" 14. és a "rövid" 21.
kromoszómát örökli, ő egészséges
emberré fejlődik, ám utódai között
ugyanúgy megjelenhetnek Down-kóros gyermekek.
Amennyiben a két vad típusú kromoszómát
örökli a szülőtől, ez a mutáció
elveszett, így utódai egészségesek
lesznek. (A kép innen származik.)
A kromoszómák inverziós mutációi sem
feltétlenül okoznak durva elváltozást. Például
Simola K. és munkatársai 1977–es cikkükben
két családot vizsgáltak meg, amelyek mindketten
egy-egy nagy méretű inverziót hordoznak 11.
kromoszómán. Az első inverziót az első
család három tagja hordozta, a második inverziót
pedig a második család nyolc tagja, ám semmilyen
kóros eltérést nem tapasztaltak egyiküknél
sem. Bár az inverziós mutációk sokszor
nem ütköznek ki rögtön, egy fontos hatásuk
mégiscsak van: ivarsejtképzéskor, a kromoszómák
átkereszteződésekor zavarokat okozhatnak, hiszen
ilyenkor nem azonos gének kerülnek egymás
közelébe, az utódokban gyakran deléciók
és duplikációk keletkeznek. Ezek a
kromoszómaátrendeződések homozigóta
formában semmilyen hátránnyal sem járnak,
ekkor az ivarsejtképzés is akadálytalanul
zajlik.
A
duplikációk azért különösen
érdekesek, mivel ilyen esetekben egy DNS szakasz
megkettőződik. Ilyenkor a megkettőződött
gének gyakorlatilag felszabadulnak a szelekciós nyomás
alól, hiszen szabadon gyűjthetnek mutációkat,
ha egy génből két példánnyal is
rendelkezik egy szervezet, az egyiknek elégnek kell lennie az
élethez, hiszen a duplikáció előtt is csak
egy példánnyal rendelkezett, ha a másik
elromlik, semmivel sem csökken az életképessége.
Sőt, amennyiben esetleg a fölös példányban
meglévő gének károsak (Mint például
a Down-kórt okozó kromoszómaduplikáció
esetén), éppen arra hat szelekciós nyomás,
hogy az egyik példány minél előbb
megváltozzon valamilyen mutációs esemény
nyomán. Ez a magyarázata annak is, hogyan
keletkezhetnek új gének a régebbiek
módosulásával, úgy, hogy az egyed
életképessége ne romoljon. Egy duplikációval
keletkezett gént akármilyen mutációs
esemény érhet, akármilyen változást
elszenvedhet, attól az eredeti példány ép,
működőképes marad. Ezek a gének
innentől kezdve gyakorlatilag szabadon változnak, ha már
egyszer működésképtelenné váltak,
a további mutációik egy amúgy is
működésképtelen génben történnek.
Ráadásul sorozatos duplikációkkal a genom
mérete is növekedhet.
Ebben az
írásban is gyakran emlegettem a szelekciós
nyomás fogalmát. De mi is ez tulajdonképpen?
Szelekciós nyomásról akkor beszélünk,
ha a különböző tulajdonságokat hordozó
egyedek nem egyforma eséllyel élnek túl és
szaporodnak. Például az emberi, X-kromoszómához
kötött vérzékenység génjére
negatív szelekciós nyomás hat, mivel az ezt
hordozó férfiak általában fiatalon
meghalnak, így egy hordozó nő fiúgyermekeinek
a fele elveszik számára, azaz kisebb eséllyel
szaporodik. Ez a negatív szelekciós nyomás, az
adott allél gyakoriságának a csökkenése
felé hat. Hogy a fenti példáknál
maradjunk, a HIV vírus elleni védettségre
pozitív szelekciós nyomás hat, hiszen az ezt
öröklő emberek kisebb eséllyel fertőződnek
meg és így ritkábban halnak meg AIDS-ben, mint
vad típusú társaik, azaz jobban szaporodnak, így
a pozitív szelekciós nyomás ez esetben az adott
allél elterjedését segíti. A szelekciós
nyomásban is meg szokás különböztetni
"természetes" és "mesterséges"
szelekciót. Ez teljességgel hibás hozzáállás,
a két folyamat semmiben sem különbözik
egymástól, a mesterséges szelekció annyit
jelent, hogy az állattenyésztő ember dönti el
a saját szempontjai szerint, hogy melyik egyed élhet és
szaporodhat tovább, ebben az esetben az emberi akarat az adott
populáció "környezete", amelyhez
alkalmazkodnia kell. Egy hasonló kísérletről
már volt szó ebben a blogban is,
ám a háziasítás minden esetben
hasonlóképpen történik. Viszont be kell,
hogy lássuk, hogy ez a szelekciós nyomás
semmiben sem különbözik attól, ami a
természetben éri az egyes populációkat,
egyszerűen bizonyos egyedek elpusztulnak, bizonyos egyedek
életben maradnak és szaporodnak.
Végül
pedig pár szót egy gyakran előkerülő
kijelentésről, miszerint a legtöbb mutáció
káros, csak ritkán találni hasznosakat. Ez
valóban igaz, mivel a mutációk véletlen
folyamatok termékei, a frissen keletkezett mutációk
túlnyomó többsége káros egy adott
környezetben, csak töredékük hasznos. Viszont a
természetes szelekció rövid úton
gondoskodik a kevésé életképes egyedek
elhalálozásáról, így a hátrányt
okozó mutációk viszonylag gyorsan eltűnnek,
míg a valamilyen formában előnyös mutációk
megmaradnak és elterjednek, mivel hordozóik nagyobb
eséllyel szaporodnak.
http://genetika.bio.u-szeged.hu/GenBS/genBS1.html
http://genetika.bio.u-szeged.hu/GenBS/Genmut.ppt
http://genetika.bio.u-szeged.hu/GenBS/genmutMech.ppt
http://genetika.bio.u-szeged.hu/GenBS/KrommutI.ppt
http://genetika.bio.u-szeged.hu/GenBS/Kromszammut.ppt
Simola K.,
Karli P., De La Chapelle A. (1977): Two pericentric inversions of
human chromosome 11. Journal of Medical Genetics 14(5): 371–374.
Köszi, ez jólesett 🙂
Jó lenne egy hasonlóan részletes az egyedfejlődésről is.
fuhur:
Egészen pontosan mi érdekelne az egyedfejlődéssel kapcsolatban? Csak mert elég nagy anyag.
Az én kisfiam transzlokációs down-szindrómás (a karyotypus 46, XY, t 14/21). Születése után rajtunk, a szülőkön elvégezték a kromoszóma vizsgálatot. A vizsgálat alapján, egyikőnk sem hordozza a mutációt.
1. Mennyi lehet az esélye egy ilyen spontán mutációnak.
2. Lehet e a vizsgálat eredménye téves (véből mutatták ki).
3. Okozhatja-e a transzlokációs down-szindrómát más öröklődő hatás.
3. Az egészséges lányom utódait érintheti-e.
KÁ:
Előrebocsájtanám, hogy nem vagyok orvos, tehát a véleményemet kellő óvatossággal kell kezelni.
1. A spontán mutáció esélyét nehéz megadni, nem nagyon találtam rá adatot, az összes Down-szindrómás beteg 2-3% -a a transzlokációs mutáns.
2. Erről egy olyan orvost kellene megkérdezni, akinek ezzel kapcsolatban gyakorlati tapasztalatai is vannak. Elméletileg nem. Ha esetleg felmerül a gyanú, gondolom újra elvégeztethetitek a vizsgálatot, elvégre veszélytelen.
3. Nem igazán értem a kérdést. A klasszikus Down-kór a 21. kromoszóma duplikációja. E mellett okozhatja a kromoszómán belül egy rövidebb szakasz duplikációja is, illetve a transzlokáció során bármelyik kromoszóma érintett lehet. De ez esetben a gyereknél pontosan tudni, milyen mutáció történt, hiszen megtalálták a 21. kromoszóma fölös példányban lévő darabját a 14. kromoszómához forrva.
4. Ha a ti kromoszómáitok normálisak, mint ahogy ezt írod, a mutáció valamelyik ivarsejtben történhetett, vagy a megtermékenyített petesejt valamelyik osztódásakor. Az sem lehetetlen, hogy a gyerek mozaikos. Erről mondtak valamit? Az egészséges lányod, vagy a ti további utódaitokat nem érintheti.
A 2. 3. kérdéshez még érdemes annyit hozzátenni, hogy a humángenetika gyakorlatilag a kivételek tudománya. Mindig van olyan beteg, akinek eddig ismeretlen baja van, sajnos a genetikai rendellenességek végtelen számúak lehetnek. Ebben az esetben viszont ilyesmiről szó sincs, a kisfiadban pontosan tudni, hol történt a mutáció, a kromoszómaképlete mindezt megmutatja. Ilyen esetben az orvos tudja mit kell keresni, azaz bármelyik szülőnél 14/21 transzlokációt, gyakorlatilag nincs esély rá, hogy elrontsa, ott a minta a kisfiad kromoszómaképletében, pontosan ezt kell megtalálnia a szülők genomjában is.
Kedves SexComb!
Köszönöm a válaszokat!
Egyébként nem mozaikos.
no akkor csavarjunk egy kicsit e témán!
Amit leirtál az OK, s végeredményben az evolúció kulcsa az, hogy a csirasejtekben létrejött mutáció hatására az utód valamennyire /valamiben eltér a szülőktől! S ha ez számára előnyös, akkor segiti mind a populáción belüli érvényesülését, mind a fennmaradását.
Feltehető viszont a kérdés:
1. a mutagén hatások a populáció egyetlen egyedét vagy többet/mindet érhetik és hoznak létre változást az utdódaik egy részében?
2. egy bizonyos mutagén hatás ( és rereményeként az a bizonyos mutáció-tipus) csak egyszer érheti az evolúció folyamán a generációs vonalat, vagy többször is?
3. ha igen, honnan lehet tudni, hogy mikor következetat be s milyen sorrendben?
primavis:
1. Egyetlen élőlény sem védett a mutációktól. Mutációk minden populáció minden egyedében történnek. Az esetleges mutagén hatások nyilván azokat az egyedeket érintik, amelyek kapcsolatba kerülnek velük.
2. Ha elolvasod a fenti példákat, látni fogod, hogy nemcsak egy adott egyed, hanem egy adott mutáns allél is újra mutálhat akárhányszor. Kellő idő alatt akárhány mutációs esemény történhet egy leszármazási vonalban.
3. Fogós kérdés. Az ecetmuslicás példákban tiszta a kép, egy törzsbe állított mutáns újabb mutációja esetén a sorrend nem kétséges, a wch allélt okozó mutációs esemény a w1 allélt létrehozó után következett be, mert a törzsbe állított w1 allélban történt. Egy természetben megfigyelt mutáció esetén sokkal homályosabb a kép. Néha a sorrend kikövetkeztethető. Például ha egy gén duplikálódott és az egyik példányban egy pontmutáció történt valamikor, nyilvánvalóan a duplikáció a korábbi mutációs esemény, a pontmutáció pedig a későbbi, mert a duplikáció előtt nem volt ami mutáljon. A többi esetben nyomra vezethet a mutáció elterjedtsége. Ha egy mutáció két leszármazási vonalban is megvan, nyilván a különválásuk előtt kellett, hogy keletkezzen, míg ha csak az egyikben, akkor inkább valószínű, hogy a különválás után keletkezett.
nem véletlenül tettem fel ezeket a kérdéseket, ugyanis a válaszokat , bár nem vagyok genetikus, szintén ismerem rájuk, de akárcsak az evolúció alapfogalmainál (változékonyság, alkalmazkodás, stb) itt is egyfajta mechanikus értelmezés kezd eluralkodni. Ennek eredményeképpen feltételezéseket tételként, bizonyitott tényként tálalnak. Ez pl. számomra a morfológiai alapokon nyugvó rendszertan átirási kisérleteinál jelent problémát s no meg, a különböző fejlődési vonalak evolúciójának a magyarázatánál. Mivel paleontológus vagyok, természetesen a rég kihalt fajok, s egyéb taxonokról van szó,a melyekből teljes DNS-t kivonni lehetetlen, ergo azt vizsgálni és összehasonlitani sem lehet, ha pedig nem lehet, akkor ennek alapján kijelentéseket sem lehet tenni, elméleteket sem lehet gyártani, ugyebár?
primavis:
“Ennek eredményeképpen feltételezéseket tételként, bizonyitott tényként tálalnak.”
No most szerény véleményem szerint ebben az írásban semmilyen feltételezést nem fejtettem ki. Egyszerűen a mutációról mint olyanról írtam, mert a korábbi írásaimhoz érkezett hozzászólásokból az derült ki, hogy maga a “mutáció” fogalma sem tiszta sokaknak. Egyszerűen csak annyit akartam megvilágítani, hogy a biológiai megfigyelések nem követik az X-men sorozatot, azaz olyan mutációt még sohasem figyeltek meg, amikor bárkinek is fémpengék nőttek volna ki a kezéből.
“Mivel paleontológus vagyok, természetesen a rég kihalt fajok, s egyéb taxonokról van szó,a melyekből teljes DNS-t kivonni lehetetlen, ergo azt vizsgálni és összehasonlitani sem lehet, ha pedig nem lehet, akkor ennek alapján kijelentéseket sem lehet tenni, elméleteket sem lehet gyártani, ugyebár?”
Kihalt fajok DNSéről ha jól olvasom egyetlen szó sem esett ebben a cikkben, ebből természetesen következtetéseket sem vontam le, elméleteket nem bontottam ki, így maga a kérdésfelvetés érdekes.
Egyébként ha már alaptalan feltételezésekről szólunk, mit szólsz az egyes számú tételhez, miszerint a mutációk csak szűk változékonyság kialakítására képesek? Erre milyen bizonyítékot hoztak a kiötlői?
nem kell védekezz, mert nem is akartalak támadni!-:)))
egyszerüen jeleztem egy jelenséget, ami még megoldásra vár. Nyügösségem oka, az erőltetett rokonsági/leszármazási hipotézisek elburjánzása.