
 A képen Janus látható, a genfi Természettudományi Múzeum teknősbékája, aki a napokban ünnepelte tizedik szülinapját. Egészsége remek, s mivel mindkét feje teljesen funkcionális, már-már a népmesékben felbukkanó sokfejű sárkányok hús-vér prototípusának tekinthető.
A képen Janus látható, a genfi Természettudományi Múzeum teknősbékája, aki a napokban ünnepelte tizedik szülinapját. Egészsége remek, s mivel mindkét feje teljesen funkcionális, már-már a népmesékben felbukkanó sokfejű sárkányok hús-vér prototípusának tekinthető.
A két darab, működő fej persze ezernyi érdekes kérdést vet fel (hogy dönti el az állat – vagy állatok, nézőpont kérdése, hány teknősről beszélünk egy helyen – melyik fej eszik, vagy melyik idegrendszer irányítja a végtagok izmait, s ha mindkettő, mi van amikor mást akarnak, vagy képes-e a két idegrendszer külön-külön aludni), de egy fejlődésbiológus számára a legizgalmasabb az, hogy miként jött létre ez a kis élőlény.
S történetesen ez az a kérdés, ahol nagy valószínűséggel százból min. 99 kollegának ugyanaz jutna az eszébe: Hans Spemann és Hilde Mangold legendás kísérlete. Spemann és Mangold kétéltű (közelebbről szalamandra) embriókat használt fel, hogy bebizonyítsa az embrió bizonyos sejtjeinek induktív tulajdonságai vannak, amelyek révén a környezetükben levő sejtekkel együtt képesek egy törzs és fej kialakítására.
 A klasszikus kísérletet később egy másik kétéltű faj, az afrikai karmosbéka (Xenopus laevis) embrióin ismételték meg számtalanszor, miközben az említett induktív tulajdonság molekuláris mibenlétét kutatták. De, hogy pontosabbann miről is van szó: a fejlődés első néhány osztódása során az embrióból fokozatosan egy többsejtből álló gömb alakul ki, amelynek egy idő után a belseje nem tömör hanem üreges lesz (ez az elsődleges bélüreg, vagy blastocoel, az embriót pedig ekkor blastulának nevezik). Ugyan a fejlődés ezen pontján kívülről úgy tűnhet, hogy az embrió kör-körösen szimmetrikus, valójában már minden későbbi testtengelye definiálva van, molekuláris szinten legalább is.
A klasszikus kísérletet később egy másik kétéltű faj, az afrikai karmosbéka (Xenopus laevis) embrióin ismételték meg számtalanszor, miközben az említett induktív tulajdonság molekuláris mibenlétét kutatták. De, hogy pontosabbann miről is van szó: a fejlődés első néhány osztódása során az embrióból fokozatosan egy többsejtből álló gömb alakul ki, amelynek egy idő után a belseje nem tömör hanem üreges lesz (ez az elsődleges bélüreg, vagy blastocoel, az embriót pedig ekkor blastulának nevezik). Ugyan a fejlődés ezen pontján kívülről úgy tűnhet, hogy az embrió kör-körösen szimmetrikus, valójában már minden későbbi testtengelye definiálva van, molekuláris szinten legalább is.
 Az embrió későbbi dorsális (háti) részén levő sejtek kis csoportja alkotja a Spemannék által leírt induktív sejtcsoportot (ezt ma Spemann organizátornak nevezik). Ha ezeket kimetszük és egy másik, blastula fázisú embrió ventrális (hasi) oldalára ültetjük be, akkor a létrejövő kis lénynek két testtengelye lesz, amelyek a faroktájon összenőnek. Ha pedig nem pont az organizátorral ellentétes oldalra kerül a transzplant, akkor még hamarabb – így el lehet jutni olyan „sziámi ikrekhez”, akik a nyaki régióban nőnek össze. Spemannék kísérlete után majdnem hat évtizednek kellett letelnie, amíg a molekuláris biológia arra a szintre jutott, hogy vizsgálni lehetett, milyen gének felelősek a jelenségért, de amikor végül sikerült az áttörés, akkor hirtelen egy rendkívül izgalmas világ tárult a kutatók elé. Az elsőnek izolált goosecoid (gsc) gén, illetve az őt követő noggin, follistatin és chordin mind hasonlónak bizonyultak atekintetben, hogy a gént egy-egy embrió ventrális oldalán kifejeztetve, mindig többé-kevésébé teljes új embrionális tengelyeket indukáltak – akárcsak Spemann transzplantátuma.
Az embrió későbbi dorsális (háti) részén levő sejtek kis csoportja alkotja a Spemannék által leírt induktív sejtcsoportot (ezt ma Spemann organizátornak nevezik). Ha ezeket kimetszük és egy másik, blastula fázisú embrió ventrális (hasi) oldalára ültetjük be, akkor a létrejövő kis lénynek két testtengelye lesz, amelyek a faroktájon összenőnek. Ha pedig nem pont az organizátorral ellentétes oldalra kerül a transzplant, akkor még hamarabb – így el lehet jutni olyan „sziámi ikrekhez”, akik a nyaki régióban nőnek össze. Spemannék kísérlete után majdnem hat évtizednek kellett letelnie, amíg a molekuláris biológia arra a szintre jutott, hogy vizsgálni lehetett, milyen gének felelősek a jelenségért, de amikor végül sikerült az áttörés, akkor hirtelen egy rendkívül izgalmas világ tárult a kutatók elé. Az elsőnek izolált goosecoid (gsc) gén, illetve az őt követő noggin, follistatin és chordin mind hasonlónak bizonyultak atekintetben, hogy a gént egy-egy embrió ventrális oldalán kifejeztetve, mindig többé-kevésébé teljes új embrionális tengelyeket indukáltak – akárcsak Spemann transzplantátuma.
Mindezt összevetve azzal, hogy azokban az embriókban, amelyekben elroncsolták a Spemann-organizátort, egyáltalán nem alakult ki fej és idegrendszer, a magyarázat egyszerűnek tűnhet: az említett géneknek mind-mind azért fontosak, hogy indukálják az idegszövet kialakulását, és hiányukban az embrió kültakarót (epidermist) hoz létre azokból a sejtekből is, amelyekből a fej alakulna ki. (A kültakaró és az idegrendszer ugyanabból a csíralemezből, az ektodermából jön létre)
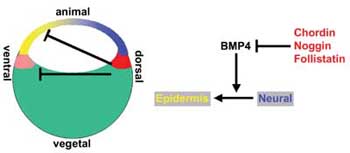 Ez ma a tudományterület többé-kevésbé elfogadott kánonja, egy fontos kivétellel: ma már nem azt gondoljuk, hogy a chordin és társai indukálják az idegrendszer kialakulását, hanem azt, hogy ők gátolják meg, hogy az epidermisben jelen levő faktorok meggátolják azt. Ugyanis, ha pl. ektodermális sejteket külön-külön nevelünk, akkor azok idegi sejtekké válnak; csak akkor lesz kültakaró belőlük, ha együtt vannak. Ez az általuk termelt BMP (Bone Morphogenetic Protein) fehérjéknek köszönhető, és mint későbbi vizsgálatok bebizonyították, az organiztátorból izolált gének döntő többsége ilyen vagy olyan formában, a BMP faktorok hatásmechanizmusát gátolja. Magyarán az ektodermális sejtek alapvető sorsa az idegrendszerré válás (ez az ún. „neural default model”, de hogy ez meg is valósuljon a BMP-k által kialakított gátlás gátlására van szükség.
Ez ma a tudományterület többé-kevésbé elfogadott kánonja, egy fontos kivétellel: ma már nem azt gondoljuk, hogy a chordin és társai indukálják az idegrendszer kialakulását, hanem azt, hogy ők gátolják meg, hogy az epidermisben jelen levő faktorok meggátolják azt. Ugyanis, ha pl. ektodermális sejteket külön-külön nevelünk, akkor azok idegi sejtekké válnak; csak akkor lesz kültakaró belőlük, ha együtt vannak. Ez az általuk termelt BMP (Bone Morphogenetic Protein) fehérjéknek köszönhető, és mint későbbi vizsgálatok bebizonyították, az organiztátorból izolált gének döntő többsége ilyen vagy olyan formában, a BMP faktorok hatásmechanizmusát gátolja. Magyarán az ektodermális sejtek alapvető sorsa az idegrendszerré válás (ez az ún. „neural default model”, de hogy ez meg is valósuljon a BMP-k által kialakított gátlás gátlására van szükség.
S hogy mindez miképpen magyarázza meg Janus esetét? Természetesen nincs szó semmilyen kísérletes transzplantációról, a természetes folyamat siklott ki. Valószínűleg (bár biztosat egy kifejlett egyedről nehéz mondani) valamilyen okból kifolyólag a fejlődés egy korai szakaszában az organizátor két részre vált szét, és ezután mindkettő tette a dolgát: létrehozott egy-egy fejet. Hogy végül ezzel együtt egy teljesen életképes egyed jött létre, az azonban mindenképpen figyelemreméltó (Blinky, a közelmúltban született kétfejű boci nem húzta sokáig, bár persze emlősök esetén a legtöbb fejlődési mechanizmus sokkal érzékenyebb a rendellenességekre, mint a hüllőknél).
(Az osztódó embriós anigif a mychemicalsoul.com-ról származik.)
Ez igen izgalmas tema. Nem irnatok bovebben az etologiai oldalarol?
Hyena,
mit értesz pontosan az „etológiai oldal” alatt? (Ha az ilyen kétfejű állatkák viselkedéséről általában – az szerintem nem nagyon vizsgált, egyrészt a minimális mintaszám miatt, másrészt pedig az esetleges megfigyelések nyilvánvalóan, hogy csak az adott speciális esetre vonatkoztathatóak, és mégcsak a faj lokális populációjára sem.)
Engem ennek a konkrét egyednek a viselkedése is érdekelne… pl. a bevezetőben említett „mi van, ha mást akar a két fej” probléma?? Nincs erről valami vizsgálata a Genfi múzeumnak?
Ha van is, sajnos en nem tudok rola.
emberben is van ilyen, a sziámi ikrek egyik fajtája.
TeknősBÉKA?!
@Caenorhabditis elegans: http://www.nyest.hu/hirek/helytelen-e-a-teknosbeka
🙂