
Minden szexuális úton szaporodó faj számára értelemszerűen igencsak fontos, hogy a szaporodáshoz szükséges mindkét nem képviseltesse magát egy-egy populációban. Ergo nem elhanyagolható kérdés, hogy a szex-determináció (azaz a nem-meghatározás) hogyan működik az adott fajban, mert ha gallyra megy, akkor a populáció rövid úton múlt időben találja magát. Érdekes módon e téren (kis túlzással) ahány ház annyi szokás, mert az állatvilágban rendkívül sok fajta megoldás született a nemek meghatározására. A számunkra jól ismert Y kromoszóma alapú (vagyis az Y kromoszóma jelenléte mindig hím identitást okoz) csak az egyik variáció a témára, madaraknál (az emlőssel nem rokon) nemi kromoszómák épp fordítva működnek (azaz ZZ kombináció hímet, míg ZW nőstényt definiál), Drosophila esetében egyszerűen az X és Y kromoszómák aránya számít, egyes halfajoknál a nemi jellegek az élet során megváltozhatnak, sok hüllő esetében a tojásokat kikeltető homok hőmérséklete határozza meg a fejlődő embrió nemét (van is így gondjuk a klímaváltozás apropóján) és a sor még folytatható lenne.

1. Ábra: A humán X kromoszóma rövidebb karja erős homológiát mutat az 1. csirke kromoszómával (ehhez szükség van azonban a legalul szemléltetett átrendeződésre – az 1. csirke kromoszóma gyűrűvé zárul, majd egy új helyen eltörik), míg a hosszabbik kar a 4. csirke kromoszómával mutat hasonlóságot (bár számos inverzió figyelhető meg). (Ahol balról jobbra lejt a szekvenciák közötti megfeleltetés grafikonja, ott azonos irányban „fut” a szekvencia, ahol pedig jobbról balra, ott ellenkező irányba, azaz valahol egy inverzió következett be). [1]
S bár az említett példák mindegyike megér egy misét, most kivételesen a bennünket legjobban érintővel foglalkoznék, pontosabban annak a sztorinak is csak a felével, az X kromoszómával. Utóbbi nemcsak azért érdekes, mert a hím emlősök sejtjeiben egy van belőle, hanem azért is mert a női sejtekben hiába van kettő, csak az egyik működik, s hogy melyik az (jelen tudásunk szerint) egy jobbára véletlen folyamat eredménye. Az X kromoszómának tavaly márciusban lett kész a térképe [1], amely számos érdekességre derített fényt. Bár a kromoszóma csak az emberi genom kb. 4%-át tartalmazza (durván 1100 gént és számos nem kódoló RNS-t), mégis a genetikailag öröklött betegségek közel 10%-a hozható vele kapcsolatba. Ennek az az oka, hogy mivel az Y kromoszómán csak az X génjeinek szűkös töredéke található, az X-en lelhető számos génre nézve a férfiak szervezete nem diploid, vagyis a génekből csak egyetlen kópia van a sejtben (a többi kromoszómánkból egy-egy pár van, így az azokon levő minden gén 2x fordul elő). Így ha az elromlik, annak meg is lesz a hatása, hiszen még véletlenül sincs hogyan kompenzálni a hibát.
A térkép továbbá igen jól bizonyítja a nemi kromoszómák autoszómális eredetét, azaz valamikor az emlősök őseinek X és Y kromoszómája egyáltalán nem volt annyira különböző mint napjainkban, hanem éppen olyan egymással szinte identikus párost alkottak mint a maradék 22 pár kromoszóma bármelyike (és ebből kifolyólag akkoriban nem is lehetett sok közük a nemi identitás kialakításához). Azonban az idők folyamán elsőrorban az Y kromoszómán lejátszódó genomi átrendeződések hatására, a két kromoszóma egre inkább elkezdett különbözni és emiatt nagy ütemben elvesztették rekombinálódási képességüket. Az autoszómális eredetet jól szemlélteti, hogy az X kromoszóma két karja igen szépen illeszthető az 1. és 4. csirke autoszómával (nem nemi kromoszómával) (1. Ábra), valamint az is, hogy az X és az Y rövid karján még ma is felfedezhetőek olyan homológ régiók amelyek azonos géneket kódolnak. Ezeket a rövid szakaszokat „pszeudoautoszómális régióknak” nevezzük, mert értelemszerűen az itt található génekből mégiscsak kettő van, vagyis úgy viselkednek mint a nem nemi kromoszómák génjei.
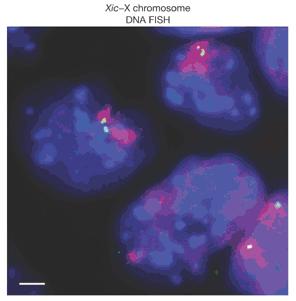
2. Ábra: Az X-kromoszóma inaktiváció során a női sejtek két X kromoszómája a XIC szekvenciák mentén rövid időre „összesimul”. (A kék folt a sejtmag, a lila paca az X kromoszómát jelöli és a zöldes-sárgás pöttyök a XIC-t.) [2]
Fentebb már utaltam röviden az X kromoszómához köthető egyik érdekes jelenségre, a női sejtekben megfigyelhető ún. X-kromoszóma inaktivációra. Ennek a véletlenszerű folyamatnak az eredményeként a két X kromoszóma egyike „kikapcsolódik”, így a hím sejtekhez hasonlóan (és a többi kromoszómától eltérően) a női sejtekben is az X kromoszómán levő génekből csak egyetlen kópia íródik át. Az inaktiváció folyamatában a kromoszóma egy speciális régiója az X-inactivation center (XIC) játszik benne kulcsszerepet, egyebek mellett itt található a Xist (X-(inactive) specific transcript) nem kódoló RNS szekvenciája. A Xist transzkriptumok az inaktiváció során a kromoszómát szabályosan beborítják, így mintegy fizikailag is lehetetlenné téve, hogy róla gének íródjanak át. De a legfrissebb jelek szerint a XIC szerepe nem merül ki ennyiben. Szinte teljesen egyszerre két külön csoport is arra a felismerésre jutott, hogy az inaktiváció során, a sejtben lelhető két X kromoszóma egymás mellé igazodik (2. Ábra), és valahogy ezen folyamat során játszák le, hogy melyikük működik tovább és melyikük vonja ki magát a forgalomból [2,3]. A jelenség igen érdekes és valószínűleg közelebb visz ahhoz, hogy megértsük az inaktiváció eleddig kicsit misztikus mechanizmusát.
És legvégül, hogy teljes legyen a kép, még egy érdekes adat: közelebbi vizsgálatok szerint mint számos biológiai jelenségre, az inaktivációra is vannak kivételek. Az X kromoszóma génjeinek kb 15%-a nem mindig kerül inaktiválásra [4]. Logikusnak tűnne, hogy a pszeudoautoszómális génekről legyen szó, de mint kiderül nemcsak ezek esnek ebbe a kategóriába. Érdekes lesz majd megérteni, hogy pontosan miért nem kapcsolódnak ki ezek a gének, és milyen hatása van a női szervezetre, hogy egyes gének transzkriptumaiból kétszer annyit hordoz sejtjeiben, mint a hímek.
[1] Ross et al. (2005) The DNA sequence of the human X chromosome. Nature 434: 3253-37.
[2] Bacher CP, Guggiari M, Brors B, Augui S, Clerc P, Avner P, Eils R, Heard E. (2006) Transient colocalization of X-inactivation centres accompanies the initiation of X inactivation. Nat Cell Biol. DOI: 10.1038/ncb1365
[3] Xu N, Tsai CL, Lee JT. (2006) Transient Homologous Chromosome Pairing Marks the Onset of X Inactivation. Science DOI: 10.1126/science.1122984
[4] Carrel L, Willard HF. (2005) X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature 434: 400-404.