
 Az aminosavsorrend adja a fehérjék elsődleges szerkezetét, az aminosavlánc aztán különféle másodlagos struktúrákat vesz fel, mint például az alfa-hélix vagy a beta-redő, ezek aztán különféle harmadlagos szerkezetekbe rendeződnek, így alakítva ki az adott fehérje háromdimenziós térszerkezetét. E szerkezet alapvetően határozza meg a fehérje funkcióját – egy enzim aktív centrumába, egy receptorfehérje ligandkötő doménjébe, egy adott transzportfehérjéhez csak egy bizonyos szerkezettel rendelkező vegyület(csoport) képes kapcsolódni. De mi a helyzet azokkal a fehérjékkel, amiknek nincs a klasszikus értelemben vett harmadlagos szerkezetük?
Az aminosavsorrend adja a fehérjék elsődleges szerkezetét, az aminosavlánc aztán különféle másodlagos struktúrákat vesz fel, mint például az alfa-hélix vagy a beta-redő, ezek aztán különféle harmadlagos szerkezetekbe rendeződnek, így alakítva ki az adott fehérje háromdimenziós térszerkezetét. E szerkezet alapvetően határozza meg a fehérje funkcióját – egy enzim aktív centrumába, egy receptorfehérje ligandkötő doménjébe, egy adott transzportfehérjéhez csak egy bizonyos szerkezettel rendelkező vegyület(csoport) képes kapcsolódni. De mi a helyzet azokkal a fehérjékkel, amiknek nincs a klasszikus értelemben vett harmadlagos szerkezetük?
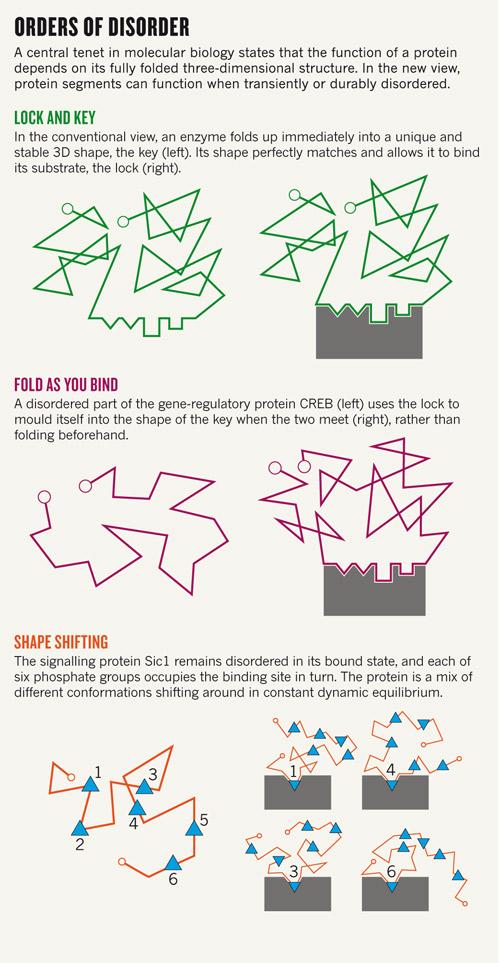
Jó néhány éve volt már, hogy Dénessel órákon át ültünk a spektrofotométer előtt, és a hődenaturált citrát szintáz fényszórásának változását mértük két általa izolált, chaperonnak gondolt rendezetlen fehérje jelenlétében. Végül többféle méréssel is sikerült igazolnunk, hogy az ERD10 és ERD14 nevű fehérjék ténylegesen dajkafehérjeként működnek, vagyis más fehérjéknek segítenek abban, hogy a térszerkezetüket felvegyék illetve megőrizzék. E két fehérje a rendezetlen fehérjék közé tartozik, teljesen funkcionális állapotukban is rendezetlenek vagyis nincs stabil, jól definiálható harmadlagos szerkezetük (IDPs – intrinsically disordered proteins). Ezek struktúrálisan a globuláris fehérjék denaturált állapotaira emlékeztetnek [1]. Azóta egyre több, akár eddig is ismert proteinről derül ki, hogy nincsen meghatározható térszerkezete, sőt a másodlagos struktúrákban sem bővelkedik. A humán gének 40%-a kódol olyan fehérjét, amelynek legalább az egyik doménje, míg negyedük olyat, amely teljesen egészében rendezetlen [2]. Többségük igencsak “befolyásos” protein (mint például a p53, BRCA1, CREB, RNS-polimeráz II, illetve az alfa-szinuklein), általában a jelátvitelben, a transzkripcióban vagy a sejtciklus szabályozásában vesznek részt.
A rendezetlenség hátterében az áll, hogy az ilyen fehérjéket (vagy fehérje alegységeket) alkotó aminosavak (Ala, Arg, Gly, Gln, Ser, Pro, Glu és Lys) nagy nettó töltést és alacsony hidrofóbicitást biztosítanak a molekulának, ami miatt az nem tud kompakt, jól definiált térszerkezetet felvenni. Különböző fizikai illetve mikroszkópos (NMR, SAXS, AMF, smFRET) mérések alapján azonban ezek a fehérjék nem állandóan rendezetlenek, hanem különböző, funkcióhoz kötött átmeneti rövid- illetve hosszútávú szerkezetekkel rendelkeznek. Egyik szerkezet a másikba például egy másik molekulával történő kapcsolódás hatására alakul át. A rendezetlenségnek köszönhetően ezek a fehérjék nagyobb specifitással rendelkeznek (ami viszont nem jár a kötés erősségének növekedésével), gyorsabban képesek e specifikus kapcsolatok kialakítására, illetve több funkciót is képesek ellátni [3].
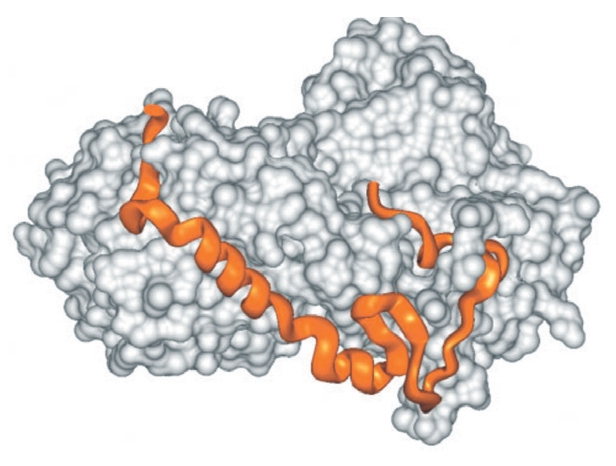
Ha a rendezetlen fehérjék létét nem is, de a működésük mechanizmusát sok kutató igencsak kételkedve fogadja. A klasszikus kulcs-zár elmélet ma már elfogad némi “lötyögést”, sőt, az allosztéria is leír interakció hatására végbemenő szerkezetváltozást, de azért a legtöbb old school fehérje-biokémikusnak a szerkezetfelismerés és a rendezetlen struktúra nem hozható egy napon szóba: “Olyan ez, mintha egy zárat egy főtt spagettivel akarnál kinyitni” – mondja a jelenségre Joël Janin, egy Gif-sur-Yvette-i szerkezetbiológus. Hihetetlennek hangzik, de – a hasonlatnál maradva – néhány esetben a spagetti (pl a CREB, lásd az alsó ábra közepén) a zárral kölcsönhatásba lépve a zárnak megfelelő kulcs alakját veszi fel. A Sic1 spagettije viszont úgy tűnik, hogy sosem vesz fel rendes harmadlagos szerkezetet, mégis kulcsszerepet játszik a sejtciklus szabályozásában [4].
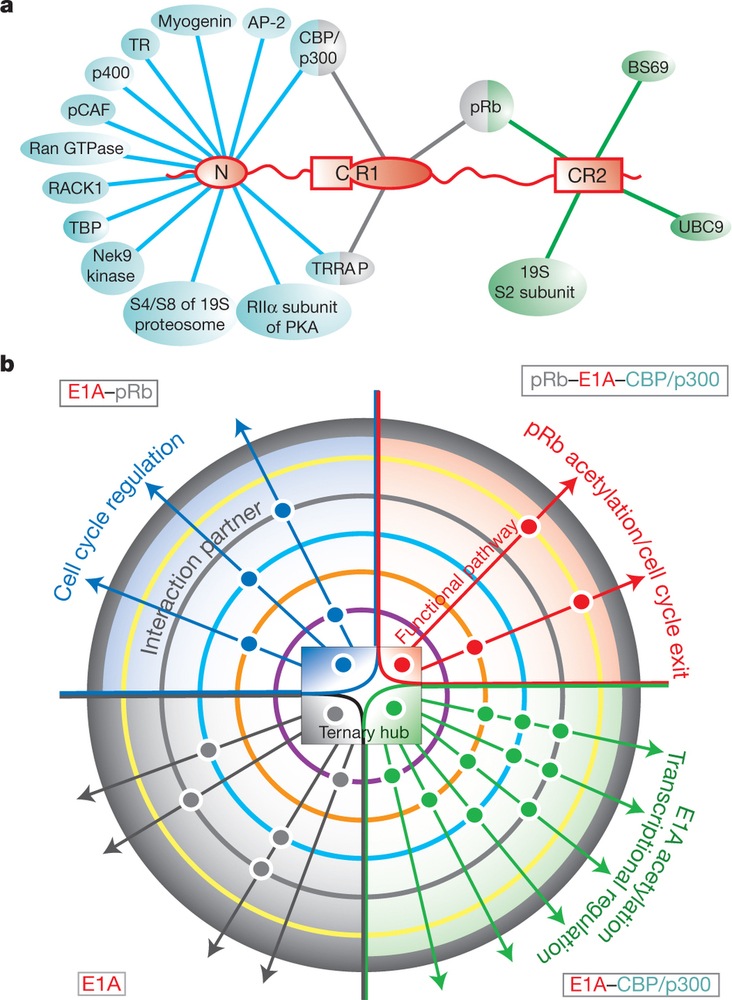
Az eukarióta sejtekben található rendezetlen p53 vagy a BRCA1 jelátviteli folyamatok ún. központi fehérjéi (“hub proteins”), mivel nagyon nagy a kapcsolati tőkéjük (számtalan partnerrel képesek komplexet alkotni). Hozzájuk hasonlatosan rendezetlen “hub” az adenovirus 1A korai fehérje (E1A) is, amely sok gazdasejtbeli szabályozófehérjével tud kapcsolatot létesíteni, ezáltal fenekestül felforgatja a fertőzött sejt jelátviteli folyamatait. A Nature legfrissebbik számában arról számol be Ferreon, hogy attól függően, milyen fehérjével lép első körben kapcsolatba az E1A, a kapcsolat által létrejött allosztérikus térszerkezet-változások miatt különböző jelátviteli utakat aktivál. Ha az elsődleges partner a CBP/p300, akkor gének átíródását befolyásolja pozitív illetve negatív irányba (lásd a kör alakú ábra jobb alsó szeletét), ha a retinoblasztóma fehérjével (pRb) képez komplexet a CBP/p300 nélkül, akkor a sejtciklust pörgeti tovább (lásd a bal felső kvadránst), ha a komplexben mind a pRb, mind a CBP/p300 jelen van, akkor sejtdifferenciációt indukál (lásd a jobb felső körcikket) [5].
Bárhogy is lássák el feladatukat, a rendezetlen fehérjék mindenképpen újraformálják a proteinek szerkezetmeghatározásán alapuló funkciófelfogást. Vagy fogalmazzunk úgy, hogy újra kéne, hogy formálják? Merthogy egyenlőre a tudományos közösség még ott sem tart, hogy a rendezetlen fehérjék bekerüljenek a tankönyvekbe.
1. http://www.matud.iif.hu/2009/09jul/08.htm.
2. Uversky, V.N. and A.K. Dunker, Understanding protein non-folding. Biochim Biophys Acta, 2010. 1804(6): p. 1231-64.
3. Tompa, P., Unstructural biology coming of age. Curr Opin Struct Biol, 2011. 21(3): p. 419-25.
4. Chouard, T., Structural biology: Breaking the protein rules. Nature, 2011. 471(7337): p. 151-3.
5. Ferreon, A.C.M., et al., Modulation of allostery by protein intrinsic disorder. Nature, 2013. 498,
: p. 390–394
Nagyon tetszik, koszi! Lehet, hogy a kovetkezo „Journal Club”-on felhasznalom!
Koch Sanyi bácsi foglalkozott egy időben sokat azzal, hogy miként találhat egymásra a vírus és az a membránfehérje, amihez kötődik. Azt mondta, hogy annak az esélye, hogy egy vírus megfertőzzön valakit, ha stabil térszerkezeteket feltételezünk a receptor és a vírus részéről is nagyjából nulla. Főleg, ha ismerjük a gyakorlatban előforduló víruskoncentrációkat (példája szerint egy poliós gyerek a városi uszodán keresztül képes ötszázat megfertőzni). Neki az volt az elképzelése, hogy mind a receptor, mind a vírus térszerkezetének dinamikusan változónak kell lennie. Erre ugyan közvetlen bizonyítéka nem volt, de már akkor is jól hangzott.
@Nádori Gergely:
Most hirtelen felindulásból rákeresve a vírus-receptor vonalra kijött egy cikk a világjárványt okozó influenzavírusokról (szintén a Dunker and Uversky szerzőpárostól),
(http://www.virologyj.com/content/6/1/69)
amiben ezt írják:
„The protein of interest is hemagglutin (HA), which is a surface glycoprotein that plays a vital role in viral entry. Distinct differences between HA proteins of the virulent and non-virulent strains are seen, especially in the region near residues 68–79 of the HA2. This region represents the tip of the stalk that is in contact with the receptor chain, HA1, and therefore likely to provide the greatest effect on the motions of the exposed portion of HA. Comparison of this region between virulent strains (1918 H1N1 and H5N1) and less virulent ones (H3N2 and 1930 H1N1) reveals that predicted disorder can be seen at this region among the more virulent strains and subtypes but is remarkably absent among the distinctly less virulent ones.”
Koch Sanyi bácsi rátapintott a lényegre.
Mi a rendezettlen fehérjék pontos definíciója? Mert itt több kategóriát meg kéne különböztetni. Először is, háromdimenziós szerkezetről akkor beszélhetünk, ha meg tudjuk mérni, többnyire kristályosítással. Azonban nem biztos, hogy amit kristályosítani lehet, annak in vivo is jól meghatározott a szerkezete, lehetséges, hogy vannak benne zsanérok, de a kristályosítás során ezek koherens módon állnak be. De még az is elképzelhető lehet, hogy valami néhány zsanérnál sokkal nagyobb mértékben rendezettlen, mégis lehet kristályosítani. Sőt az ellentettjét is el tudom képzelni, hogy valaminek van in vivo jól meghatározott szerkezete, de mégsem lehet kristályosítani. Nem tudom, hogy mi az igazság, ha esetleg ezekről az esetekről hallhatnánk egy hozzáértőtől pár szót, azt megköszönném. Aztán a kérdéses még a méretskála és az eloszlás is, mert minden fehérjében van szinte egy-két rendezettlen oldallánc, vagy hurok, de lehetnek nagyobb domének is rendezettlenek.
aminek in vivo van jól meghatározott szerkezete
@G. M. E.:
A definíció úgy szól, hogy nincs egy stabil, jól definiálható harmadlagos szerkezetük, hanem különböző, funkcióhoz kötött átmeneti szerkezetekkel rendelkeznek, amelyek leginkább a denaturált globuláris fehérjékre hasonlítanak. Vagyis a definíció szerintem a teljes hosszukban rendezetlen fehérjékre vagy egy-egy rendezetlen fehérjedoménre vonatkozik.
Lehet őket kristályosítani, de a szerkezetvizsgálatok nem egy homogén, reprodukálható eredményt adnak.
Vannak olyan módszerek, amikkel azért az in vivo 3D szerkezetre is lehet információt nyerni, kristályosítás nélkül, mint pl a cikkben is említett smFRET, és ha nem tévedek az AFM-os vizsgálatokhoz is elég csak immobilizálni a fehérjét.
Igen, a nagyságrend az fontos kérdés, mert attól, hogy egy rövid szakasz rendezetlen (egy zsanér, ahogy mondod), de attól még a fehérje kompakt és jól leírható szerkezettel bír, de ahogy a cikkben is szerepel, a humán fehérjék negyede teljes hosszában rendezetlen.
@orient expressz: Jol gondolom, hogy oldatban az NMR is valaszt adhat arra van-e allando szerkezete az adott feherjenek?
@fordulo_bogyo:
Igen,igen,az NMR-nél oldatban van a vizsgálandó fehérje, vagyis ahhoz sem kell kikristályosítani.