
Legutóbb érkezett egy kérés, hogy ha már más immunrendszerekről írunk, akkor egy rövid összefoglaló erejéig említsük meg az emlős immunrendszert is, úgyhogy az adaptív immunitás egy folyamatáról szeretnék egy rövid, zanzásított leírást adni, az immunglobulin gének átrendeződéséről.
Legutóbb érkezett egy kérés, hogy ha már más immunrendszerekről írunk, akkor egy rövid összefoglaló erejéig említsük meg az emlős immunrendszert is, úgyhogy az adaptív immunitás egy folyamatáról szeretnék egy rövid, zanzásított leírást adni, az immunglobulin gének átrendeződéséről.
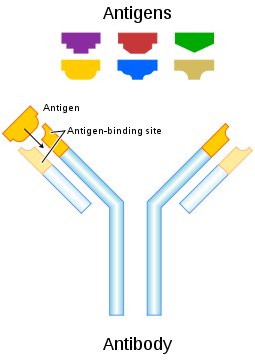 Egy immunglobulin molekula négy polipeptidláncból áll, két egyforma nehéz láncból és két egyforma könnyű láncból, ezt a négy fehérjét diszulfidhidak kapcsolják össze, így jön létre a közismert Y alakú szerkezet. Az Y két szárán található az antigénkötő zseb, a molekulának ez a szakasza alkot komplexet az általa felismert struktúrával (antigénnel). Az immunglobulin molekulákat a B-limfociták termelik, szekretálhatják oldott ellenanyagként és a membránjukhoz kötött fehérjemolekulaként is megtermelhetik, ekkor B-sejt receptor felismerőegységének is nevezik. Mire jó egyáltalán? Ha egy immunglobulin molekula komplexet képez az antigénnel, az önmagában akadályozhatja annak működését. Például egy vírus burokfehérjéhez kötődő ellenanyag megakadályozhatja, hogy a vírus bejusson a sejtbe. Egy sejtfelszíni fehérjéhez kötődő ellenanyag akadályozhatja a fehérje működését, ha az éppen egy baktérium fertőzőképességéhez fontos molekula, máris gyengíti a kórokozót. Mivel egy ellenanyag-molekula egyszerre két azonos epitópot képes kötni, megfelelő koncentrációban képes egymáshoz kötni az antigénrészecskéket, így megkönnyítve az eltávolításukat. Ezen kívül az antigénhez kötődő immunglobulin molekula fontos jel a veleszületett immunitás effektorfolyamatainak is, egy ellenanyaggal borított antigénrészecskét sokkal könnyebben kebeleznek be a fagocitasejtek, amelyeknek immunglobulinkötő receptorai vannak. Az antigén felszínéhez kötődő ellenanyag molekulák aktiválják a komplementrendszert, így beindítják a véralvadást is, ezzel mintegy csapdába ejtve a kórokozókat. Így ha a szervezet rendelkezik olyan immunglobulin molekulákkal, amelyek képesek felismerni egy kórokozót, azonnal hatékony immunválaszt képes indítani ellene.
Egy immunglobulin molekula négy polipeptidláncból áll, két egyforma nehéz láncból és két egyforma könnyű láncból, ezt a négy fehérjét diszulfidhidak kapcsolják össze, így jön létre a közismert Y alakú szerkezet. Az Y két szárán található az antigénkötő zseb, a molekulának ez a szakasza alkot komplexet az általa felismert struktúrával (antigénnel). Az immunglobulin molekulákat a B-limfociták termelik, szekretálhatják oldott ellenanyagként és a membránjukhoz kötött fehérjemolekulaként is megtermelhetik, ekkor B-sejt receptor felismerőegységének is nevezik. Mire jó egyáltalán? Ha egy immunglobulin molekula komplexet képez az antigénnel, az önmagában akadályozhatja annak működését. Például egy vírus burokfehérjéhez kötődő ellenanyag megakadályozhatja, hogy a vírus bejusson a sejtbe. Egy sejtfelszíni fehérjéhez kötődő ellenanyag akadályozhatja a fehérje működését, ha az éppen egy baktérium fertőzőképességéhez fontos molekula, máris gyengíti a kórokozót. Mivel egy ellenanyag-molekula egyszerre két azonos epitópot képes kötni, megfelelő koncentrációban képes egymáshoz kötni az antigénrészecskéket, így megkönnyítve az eltávolításukat. Ezen kívül az antigénhez kötődő immunglobulin molekula fontos jel a veleszületett immunitás effektorfolyamatainak is, egy ellenanyaggal borított antigénrészecskét sokkal könnyebben kebeleznek be a fagocitasejtek, amelyeknek immunglobulinkötő receptorai vannak. Az antigén felszínéhez kötődő ellenanyag molekulák aktiválják a komplementrendszert, így beindítják a véralvadást is, ezzel mintegy csapdába ejtve a kórokozókat. Így ha a szervezet rendelkezik olyan immunglobulin molekulákkal, amelyek képesek felismerni egy kórokozót, azonnal hatékony immunválaszt képes indítani ellene.
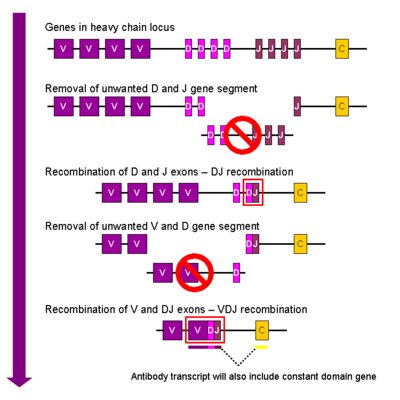 Egy immunglobulin molekula általában egyetlen epitópot ismer fel, egy érett emberi immunrendszer egy adott pillanatban akár tíz a kilencediken nagyságrendű különböző ellenanyagmolekulát képes termelni. Az könnyen belátható, hogy az emberi genomban nem férne el sokmilliónyi immunglobulin gén, mivel összesen huszonötezer fehérjekódoló génünk van. Viszont egy B-sejt csak egyetlen ellenanyagmolekulát termel, ha osztódik, a klónjai ugyanazt az immunglobulin molekulát fogják termelni, így a sokmilliónyi különböző molekulához nem szükséges a genomban sokmilliónyi gén, minden egyes B-sejt genomja egyetlen immunglobulin fehérjét kódol, csak éppen mind másikat. Nyilvánvaló kérdés, hogy akkor honnan kerülnek a B-sejtek genomjába az egymástól különböző immunglobulin gének? Szomatikus génátrendeződések hozzák létre az egyedi géneket. A csíravonalban az immunglobulin gének egészen más képet mutatnak, mint a limfocitákban. Egy lókuszban nagyon nagy számú génszegment helyezkedik el egymás mellett. A nehézlánc V, D és J szegmentekből alakul ki, egy egér genomban például a nehéz lánc lókusz 250-1000 V génszegmentet tartalmaz, 4 J szegmentet és 12 D szegmentet. A B-sejtek érése során elsőként egy véletlenszerűen kiválasztott D szegment és egy véletlenszerűen kiválasztott J szegment átrendeződése zajlik le, a köztük lévő DNS szakasz egyszerűen kivágódik, így egymás mellé kerül egy D és egy J szegment. Ezek után a DJ szegment és egy véletlenszerűen kiválasztott V szegment közti DNS szakasz vágódik ki, így alakul ki szomatikus rekombinációval az átrendezett gén, ami az immunglobulin molekula antigénkötő régióját kódolja. A könnyűlánc gének hasonlóan rendeződnek át, ott azonban nincsenek D szegmentek, csak V és J, a kappa lánc gén egérben 250 V szegmentet és 4 J szegmentet, a lambda lánc gén 2 V szegmentet és 3 J szegmentet tartalmaz. <1> Ebből könnyen kiszámolható, hogy csak a nehézlánc génszegmentekből 10-40000 különböző változat jöhet létre, a könnyű lánc esetén a kappa lánc génekből ezer, a lambda lánc génekből hat. Mivel egy immunglobulin molekula két azonos nehéz és két azonos könnyű láncból áll és gyakorlatilag bármelyik nehézlánc bármelyik könnyű lánc mellé kerülhet, csak a génszegmentek átrendeződésével tízmilliós nagyságrendű különböző immunglobulin gén alakulhat ki.
Egy immunglobulin molekula általában egyetlen epitópot ismer fel, egy érett emberi immunrendszer egy adott pillanatban akár tíz a kilencediken nagyságrendű különböző ellenanyagmolekulát képes termelni. Az könnyen belátható, hogy az emberi genomban nem férne el sokmilliónyi immunglobulin gén, mivel összesen huszonötezer fehérjekódoló génünk van. Viszont egy B-sejt csak egyetlen ellenanyagmolekulát termel, ha osztódik, a klónjai ugyanazt az immunglobulin molekulát fogják termelni, így a sokmilliónyi különböző molekulához nem szükséges a genomban sokmilliónyi gén, minden egyes B-sejt genomja egyetlen immunglobulin fehérjét kódol, csak éppen mind másikat. Nyilvánvaló kérdés, hogy akkor honnan kerülnek a B-sejtek genomjába az egymástól különböző immunglobulin gének? Szomatikus génátrendeződések hozzák létre az egyedi géneket. A csíravonalban az immunglobulin gének egészen más képet mutatnak, mint a limfocitákban. Egy lókuszban nagyon nagy számú génszegment helyezkedik el egymás mellett. A nehézlánc V, D és J szegmentekből alakul ki, egy egér genomban például a nehéz lánc lókusz 250-1000 V génszegmentet tartalmaz, 4 J szegmentet és 12 D szegmentet. A B-sejtek érése során elsőként egy véletlenszerűen kiválasztott D szegment és egy véletlenszerűen kiválasztott J szegment átrendeződése zajlik le, a köztük lévő DNS szakasz egyszerűen kivágódik, így egymás mellé kerül egy D és egy J szegment. Ezek után a DJ szegment és egy véletlenszerűen kiválasztott V szegment közti DNS szakasz vágódik ki, így alakul ki szomatikus rekombinációval az átrendezett gén, ami az immunglobulin molekula antigénkötő régióját kódolja. A könnyűlánc gének hasonlóan rendeződnek át, ott azonban nincsenek D szegmentek, csak V és J, a kappa lánc gén egérben 250 V szegmentet és 4 J szegmentet, a lambda lánc gén 2 V szegmentet és 3 J szegmentet tartalmaz. <1> Ebből könnyen kiszámolható, hogy csak a nehézlánc génszegmentekből 10-40000 különböző változat jöhet létre, a könnyű lánc esetén a kappa lánc génekből ezer, a lambda lánc génekből hat. Mivel egy immunglobulin molekula két azonos nehéz és két azonos könnyű láncból áll és gyakorlatilag bármelyik nehézlánc bármelyik könnyű lánc mellé kerülhet, csak a génszegmentek átrendeződésével tízmilliós nagyságrendű különböző immunglobulin gén alakulhat ki.
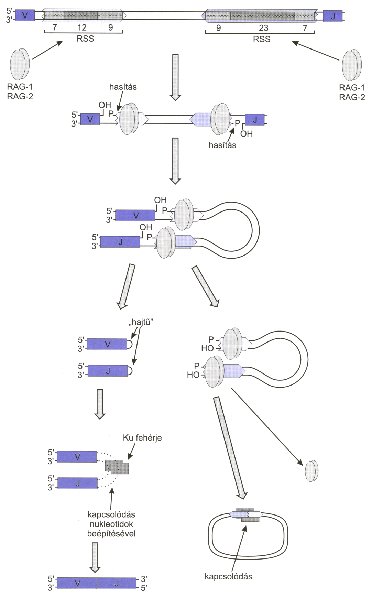 Érdemes megnézni, hogyan zajlik ez a génátrendeződés. Minden génszegment egy-egy rekombinációs szignálszekvenciát (RSS) tartalmaz az 5′ és a 3′ végén. Ezek egy hét és kilenc bázispárnyi konzervált DNS szekvenciából állnak és a köztük lévő tizenkét vagy huszonhárom bázispárnyi spacer DNS szakaszból. Ezeket az RSS szekvenciákat ismeri fel a RAG1 és a RAG2 fehérje, amelyek kettősszálú DNS törést hoznak létre a szignálszekvenciáknál, amelyek egy DNS hajtű szerkezetté záródnak. Ezen a ponton egy fehérjekomplex alakul ki a DNS végein, amelyek hasítják a hajtűstruktúrát, ekkor a terminális deoxiribonukletotid transzferáz enzim templát nélkül, véletlenszerűen bázisokat illeszt a DNS szálak végeire, majd egy DNS ligáz enzim összekapcsolja a két DNS molekulát. Így kapcsolódik össze két génszegment, ám mivel közéjük véletlenszerűen is beépülhetnek bázisok, olyan DNS szakaszt is tartalmazhat egy átrendeződött immunglobulin gén, amely nem kódolt a genomban, ez jelentősen megnöveli az ellenanyagok változékonyságát.
Érdemes megnézni, hogyan zajlik ez a génátrendeződés. Minden génszegment egy-egy rekombinációs szignálszekvenciát (RSS) tartalmaz az 5′ és a 3′ végén. Ezek egy hét és kilenc bázispárnyi konzervált DNS szekvenciából állnak és a köztük lévő tizenkét vagy huszonhárom bázispárnyi spacer DNS szakaszból. Ezeket az RSS szekvenciákat ismeri fel a RAG1 és a RAG2 fehérje, amelyek kettősszálú DNS törést hoznak létre a szignálszekvenciáknál, amelyek egy DNS hajtű szerkezetté záródnak. Ezen a ponton egy fehérjekomplex alakul ki a DNS végein, amelyek hasítják a hajtűstruktúrát, ekkor a terminális deoxiribonukletotid transzferáz enzim templát nélkül, véletlenszerűen bázisokat illeszt a DNS szálak végeire, majd egy DNS ligáz enzim összekapcsolja a két DNS molekulát. Így kapcsolódik össze két génszegment, ám mivel közéjük véletlenszerűen is beépülhetnek bázisok, olyan DNS szakaszt is tartalmazhat egy átrendeződött immunglobulin gén, amely nem kódolt a genomban, ez jelentősen megnöveli az ellenanyagok változékonyságát.
Magyarán egy B-sejt az érése során átrendezi a saját genomját, egy véletlenszerű immunglobulin molekulát kódoló gént alakít ki. Mivel ez a folyamat minden egyes B-sejtben végbemegy, minden egyes érett B-sejt egy egyedi immunglobulin molekulát termel majd. Nyilván ezen molekulák nagy része működésképtelen, korai STOP kodont tartalmaz, semmit sem ismer föl, vagy éppen olyan antigént ismer föl, amivel sohasem találkozik a szervezet, ezek nyilván fölöslegesek. Ezen kívül veszélyes lehet egy olyan ellenanyag, ami egy saját fehérjét ismer föl, hiszen azzal az immunrendszer a saját tested szöveteit károsítaná, autoimmun betegségek kialakulásához vezetne. Egy B-sejtben a génátrendeződés a csontvelőben zajlik le, amennyiben egy B-sejt itt átrendezett receptora olyan antigént ismer föl, ami a csontvelőben is jelen van, apoptózissal elpusztul. Így eleve kiszelektálódnak a szervezetben mindenütt jelen lévő saját epitópokat felismerő receptorokat hordozó sejtek. Azonban a csontvelőben nincs jelen minden antigénként működő saját struktúra, így olyan B-sejtek is megérhetnek és kijuthatnak a csontvelőből, amelyek valamilyen saját antigént felismerő ellenanyagot termelnek. Ezek nem feltétlenül okoznak betegséget, mivel ahhoz, hogy egy B-sejt aktiválódjon és elkezdjen ellenanyagot termelni, szükséges, hogy egy T-sejttől megerősítő jelet kapjon, e nélkül elmarad az aktiváció, nem termel majd ellenanyagot. Így bár nyilván rendszeresen képződnek autoreaktív ellenanyagok termelésére képes B-sejtek, ezek nem károsítják a szervezetet, mert sohasemaktiválják őket a T-sejtek. Ugyanígy azok a B-sejtek sem aktiválódnak, amelyek olyan ellenanyagot termelnének, amely éppen semmit sem ismer föl, hiszen nem kapnak jelet a T-sejtektől. Ezt a jelenséget nevezzük klónszelekciónak.
Azok a B-sejtek, amelyek olyan immunglobulin molekulát termelnek, amely felismer egy éppen jelenlévő idegen antigént, megerősítést kapnak a T-sejtektől, villámgyorsan osztódni kezdenek, így viszonylag rövid idő alatt egy sikeres klón nagy számban jelenik meg. Az osztódó B-sejtek differenciálódnak is, az effektorsejtek nagy mennyiségű ellenanyagot termelnek és juttatnak a keringésbe, míg a sejtek kis hányada memóriasejtté alakul. A memóriasejtek évtizedeken át élnek, ha nem találkoznak az általuk felismert antigénnel, nem aktiválódnak, ám amint újra kapcsolatba kerülnek az általuk felismert antigénnel, azonnal osztódni kezdenek, így ha a szervezet másodjára találkozik egy kórokozóval, már sokkal gyorsabban képes az ellenanyagválaszra, hiszen nem kell véletlenszerű átrendeződésekkel kialakítania a mikróbát felismerő receptort, már ott van készen.
Ez a nagyon zanzásított leírás csak vázlatosan mutatja be a folyamatot, de a fő szereplők bemutatására alkalmas és bemutatja az immunválasz működését: A csontvelőben naponta milliószámra differenciálódnak B-sejtek, amelyek mindegyike egy kombinatorikus génátrendeződéssel képződött egyedi immunglobulin gént hordoz. Ezek utána a keringésbe jutnak, ahol nyilván találkoznak a szervezetbe került mikróbákkal. Azok a klónok, amelyek olyan immunglobulin fehérjét képesek termelni, amely felismer valamilyen jelenlévő idegen antigént, megerősítést kapnak a T-sejtektől, aktiválódnak, osztódni kezdenek, majd nagy mennyiségű ellenanyagot termelnek. Az aktivált B-sejtpopulációból memóriasejtek is képződnek, amik évtizedeken át megőrzik a B-sejt klónt, ami egyszer sikeres immunválaszt adott. Ebből következik, hogy az adaptív immunitás nem örökölhető. Ha a szülő már egyszer sikeres immunválaszt adott mondjuk egy baktériumfertőzésre, ő maga védetté válik, hiszen ott vannak a szervezetében azok a memóriasejtek, amelyek nagyon gyors ellenanygaválaszt képesek adni, ám az utódainak ettől még ugyanúgy újra le kell győzni a kórokozót, a sikeres B-sejt klón nem adható át nekik.
<1>Gergely János, Erdei Anna: Immunbiológia, Medicina kiadó, 2000