
 A Harvardon dolgozó Arkhat Abzhanov-ot már évek óta izgatja az arckoponya evolúciója. Hogy miként alakult ki, hogyan evolválódott/evolválódik egyes fajokban (posztdokként Cliff Tabin laborjában pintycsőr evo-devoval foglalkozott). Most az Evolution című újságban megjelent cikk egy kicsit nagyobb léptékű változásnak járt utána, hogy hogyan is alakult át a más gerincesek viszonylag kisméretű praemaxilla csontja, hogy a madarak jellegzetes alakú csőrét hozhassa létre.
A Harvardon dolgozó Arkhat Abzhanov-ot már évek óta izgatja az arckoponya evolúciója. Hogy miként alakult ki, hogyan evolválódott/evolválódik egyes fajokban (posztdokként Cliff Tabin laborjában pintycsőr evo-devoval foglalkozott). Most az Evolution című újságban megjelent cikk egy kicsit nagyobb léptékű változásnak járt utána, hogy hogyan is alakult át a más gerincesek viszonylag kisméretű praemaxilla csontja, hogy a madarak jellegzetes alakú csőrét hozhassa létre.
A csőr evolúciója ugyanis jól követhetően ennek az egy bizonyos csontnak a megnyúlásáról szól és bár a csőr kialakulásának persze nyilván előfeltétele volt a fogak elvesztése (amiről már korábban írtunk itt és itt), azok hiánya önmagában édeskevés lenne az anatómiailag modern csőr kialakulásához.
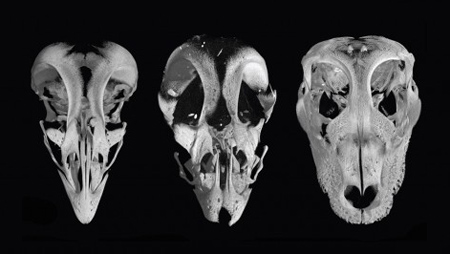
 Hogy miként is következett be a (jobb oldali ábra jobb oszlopában pirossal jelölt) csont méretváltozása, ahhoz előbb azt szükséges megértenünk, hogy különböző fajokban az arckoponya adott részének fejlődése alatt milyen gének aktívak, illetve ezen a téren találunk-e valamilyen különbséget a madarak, illetve a hüllők között.
Hogy miként is következett be a (jobb oldali ábra jobb oszlopában pirossal jelölt) csont méretváltozása, ahhoz előbb azt szükséges megértenünk, hogy különböző fajokban az arckoponya adott részének fejlődése alatt milyen gének aktívak, illetve ezen a téren találunk-e valamilyen különbséget a madarak, illetve a hüllők között.
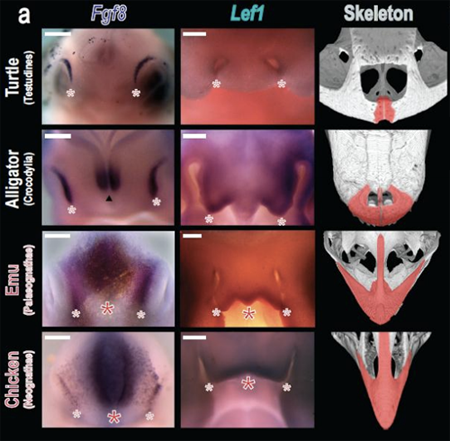
Abzhanovék két gén esetében is látványos eltérést dokumentátak: mind az FGF-jeltávitel útvonal egyik ligandja, az Fgf8, mind a Wnt jelátvitel egyik effektor transzkripciós faktora, a Lef1, sokkal kiterjedtebb expressziót mutatott az emukban és a csirkékben, mint a teknősökben, vagy az alligátorokban. A két jelátviteli útvonal rendkívül pleiotróp, vagyis kiscsillió szerepük van az egyedfejlődésben, és eddig is ismert volt a szerepük az arcfejlődés egyes aspektusaiban, és most úgy tűnik, valami újat tudtunk meg róluk.
Persze a nagy kérdés ilyenkor, hogy van-e ok-okozati összefüggés a megnövekedett Fgf8 és/vagy Lef1 expressziós domén, valamint a praemaxillák megváltozott morfológiája között. Ennek vizsgálatához olyan reagenseket használtak, amelyek az említett útvonalakat gátolják: apró golyókat áztattak ezekbe az anyagokba, majd a fejlődő embriók megfelelő pontjára helyezték és figyelték a hatást.
Ami nem maradt el: a gracilis csőr ezekben a csirkeembriókban egy sokkal robusztusabb formát öltött, elmaradt a praemaxillák madarakra oly jellemző fúziója és úgy általában is egy alligátor-szerű arcberendezés jött létre. Az alakváltozás a szájpadot felépítő palatumot is érintette, az is egy sokkal “ősibb” formát öltött (ezt az alábbi kép d. ábráján lehet megfigyelni).
Arra is fény derült, hogy az Fgf8 és Lef1 expresszió között is ok-okozati összefüggés van, hiszen az FGF-inhibítorral kezelt állatokban a Lef1 kifejeződése rendre lecsökkent a fejlődő arc középső részén (azaz pont ott, ahol a madarakban eredetileg megnőtt), vagyis az utóbbi gén működését az FGF jelátvitel szabályozza (direkt vagy indirekt módon).
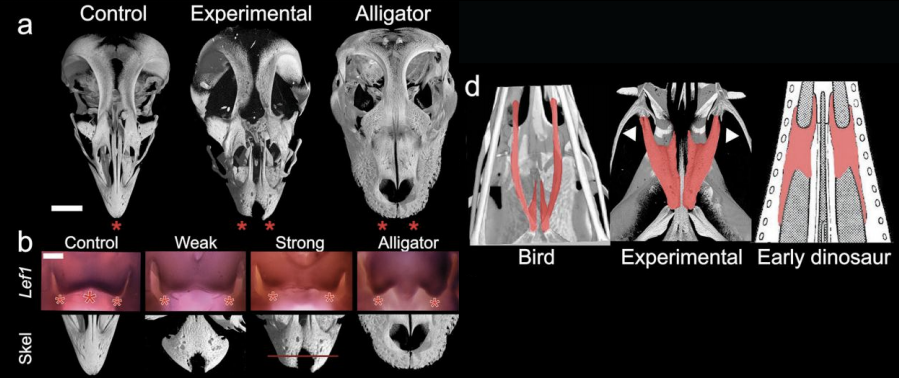
Mindezekből az a következtetés vonható le, Abzhanovék olvasatában, hogy a csőr-evolúció fontos állomása lehetett az FGF- és Wnt-útvonalak aktiválódása a fejlődő embriók leendő arcának középső részén, ami még a madarak evolúcijának hajnalán bekövetkezett. (Az emu és csirke párhuzamos vizsgálata azért mérvadó, mert a madarak két nagy alosztályát képviselik: a Paleognathae fedőnevű futómadár-alkatúakat, illetve az újmadár-szabásóakat (Neognathae). Vagyis azok a jellegek, amelyek mindkét csoportban megvannak jó eséllyel a madár-evolúció hajnalán tűnhettek fel.)
Abzhanovék munkájának azért akadnak kritikusai is: a ScienceNews-on megjelent cikk szerint Ralph Marcucio csoportja például, ha nem is tartja lényegtelennek az Abzhanovék által leírtakat, sokkal fontosabbanak gondolják a SHH (Sonic Hedgehog) gén működésében fellépő változásokat a csőr kialakulása szempontjából.
A csőr evolúciója ezzel persze nem ért véget, sőt, épp hogy elkezdődött: mint azt a Grant házaspár évtizedes munkája alaposan dokumentálta a galapagosi pintyek esetében, a folymat megállás nélkül zajlik, és ma már egyre jobban értjük azt is, hogy milyen gének játszanak szerepet a különböző csőrtípusok létrehozásában – ahogy korábban említettem pont Abzhanov korábbi munkáinak köszönhetően (pl. BMP4, CaM, ALX1).
UPDATE: Itt van egy rövid beszélgetés is, ahol az első szerző, Bhart-Anjan Bhullar beszélget Ira Flatow-val a ScienceFridayben az eredményekről.
Bhullar BA, Morris ZS, Sefton EM, Tok A, Tokita M, et al. (2015) A molecular mechanism for the origin of a key evolutionary innovation, the bird beak and palate, revealed by an integrative approach to major transitions in vertebrate history. Evolution doi: 10.1111/evo.12684.
Érdemes még elolvasni Abzhanov Node-on megjelent komment-posztját is, amiben méginkább perspektívába teszi a dolgot és ír a korábbi Nature cikkukról is (ami az én figyelmemet elkerülte):
thenode.biologists.com/how-the-bird-got-its-beak/research/
Megengedve, hogy a madárembriók egyedfejlődésének a manipulációjából le lehet vonni ilyen következtetéseket, azonban paleontológus szemmel nézve a madárcsőr evolúciója másként néz ki. A madarak diapszid koponyájú hüllőkből alakultak ki, a dinókkal, pterosauruszokkal, krokodilokkal, gyíkokkal, stb. együtt. Erre a koponyatípusra jellemző, hogy a szemgödrök összeérnek s így az az agyüreg mögöttük végződik. Ez az arckoponya csontjainak jelentős változásával is jár s szinte valamennyi típusnál meghosszabbodnak az állkapcsok s kialakulhat egy csőrszerű képződmény. Ennek a szerepe elsősorban a táplálkozásban van s ezért nem találunk nagy formagazdagságot az esetükben. Fogakkal ellátva vagy anélkül, a cél a táplálék megragadása, feldarabolása, megőrlése. Ez a helyzet a dinoszauruszok, pteroszauruszok, de a gyíkok, krokodilok, a gyíkfarkú ősmadarak esetében is. (Kivételt csak a kígyók képeznek, ahol a fogak – ha vannak, csak a préda fogva tartását és megbénítását – a méreggel bíróknál – biztosítják s emiatt a csőszerű megnyúlás is fejletlen). A valódi madarak esetében a madárcsőr kialakulásának az oka a kéz és a szárnyujjak szerkezetének és a szerepének a megváltozása. A szárnnyá alakult mellső végtag mozgásbeli funkcióján kívül, a többi egykori szerep mind a csőrre hárult. Ennek következtében alakult ki a neognath madárkoponya típus. (A különbség a paleognath és neognath koponyatípus közt az, hogy az előbbinél a basisphenoideum és a pterygoideumok közt egy őket összekötő nyúlvány van, míg az utóbbinál ez hiányzik s így a szájpadcsont és a röpcsontok mozgékonyan izesűlhetnek egymáshoz). A mai futómadarak többségénél ez a jelleg nem alakult ki, megmaradt az ősibb forma, ami arra utal hogy az evolúciójuk során korán elváltak a neognath típustól, másrészt az életmódjuk nem is tette szükségessé ennek a jellegnek a megjelenését (a mai futómadarak csőre – a szélsőségesen specializálódott kivicsőr kivételével – alig különbözik a gyíkfarkú madarakétól s egyben az Archosaurida ősökétől).