
 A növények anyagcseréje fényfüggő (valódi fényevőkről lévén szó), és ennek megfelelően számos olyan tulajdonságuk van, ami a fényfelvételt optimalizálja. Ezek egyike a fototropizmus jelensége, amely során a növények a fény felé nőnek, vagy a fény felé fordulnak (lásd napraforgó). Nem ez az egyetlen fény-függő folyamat, persze, a növények napi biológiai ritmusa és a tavaszi virágzás időpontjának meghatározása is természetesen fény-függő (utóbbi esetben a növény érzékelni tudja az egyre hosszabb nappalokat és ezt követően indítja el a virágképzés-programját). Érdekes módon a fototropizmust és a biológiai ritmusokat azonban egyáltalán nem ugyanaz a molekuláris rendszer szabályozza, már a fény érzékelése is más-más molekuláris komponensek révén következik be.
A növények anyagcseréje fényfüggő (valódi fényevőkről lévén szó), és ennek megfelelően számos olyan tulajdonságuk van, ami a fényfelvételt optimalizálja. Ezek egyike a fototropizmus jelensége, amely során a növények a fény felé nőnek, vagy a fény felé fordulnak (lásd napraforgó). Nem ez az egyetlen fény-függő folyamat, persze, a növények napi biológiai ritmusa és a tavaszi virágzás időpontjának meghatározása is természetesen fény-függő (utóbbi esetben a növény érzékelni tudja az egyre hosszabb nappalokat és ezt követően indítja el a virágképzés-programját). Érdekes módon a fototropizmust és a biológiai ritmusokat azonban egyáltalán nem ugyanaz a molekuláris rendszer szabályozza, már a fény érzékelése is más-más molekuláris komponensek révén következik be.
A fototropizmus szabályozására szofisztikált molekuláris mechanizmusok alakultak ki a növényekben, amelyek kulcsszereplője a rövidhullámhosszú kék-fényre érzékeny fototropin (Phot) fehérjék. Ez szinte minden ismert növényfajban így működik, kivételt csak a spórával szaporodó, úgynevezett kriptogám fajok képeznek, amelyek a hosszú hullámhosszú vörös fényre érzékenyek. Ez azért is érdekes, mert a többi növényben is léteznek olyan fehérjék is, amelyek erre a hullámhosszra specializálódtak, de ezek pont a napi ritmus és virágzás szabályozásában részt vevő fitokrómok (Phy). A kriptogám fajokhoz tartozó páfrányok körében azonban nem is a fitokrómok vették át a fototropin szerepét, hanem ennél izgalmasabb módon egy Phot-Phy hibrid fehérje jött létre az evolúció során, amit neokrómnak (Neo) nevezünk.
Sokáig úgy tűnt, hogy a páfrányok és néhány algafaj rendelkezik csak neokrómmal és a jelek mind arra mutatnak, hogy a két neokróm egymástól függetlenül jött létre. De hogy miként, az vált váratlanul izgalmas kérdéssé.
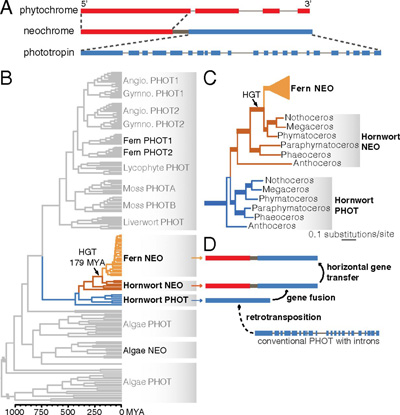 Ugyanis számos növény genomját végigfésülve váratlanul a becősmohák (angoul hornwort) genomjában is rábukkantak egy neokróm génre, és a későbbi filogenetikai vizsgálatok mind arra világítottak rá, hogy a páfrányok neokrómja nagyon hasonlít erre. Olyannyira, hogy a legésszerűbb magyarázat, hogy a páfrányok a moháktól vették á a gént.
Ugyanis számos növény genomját végigfésülve váratlanul a becősmohák (angoul hornwort) genomjában is rábukkantak egy neokróm génre, és a későbbi filogenetikai vizsgálatok mind arra világítottak rá, hogy a páfrányok neokrómja nagyon hasonlít erre. Olyannyira, hogy a legésszerűbb magyarázat, hogy a páfrányok a moháktól vették á a gént.
A páfrány-becősmoha neokróm ugyanis jellegzetes szerkezettel rendelkezik, 5′ végében hordozza a Phy szekvenciát, a 3′ végén pedig a Phot szekvencia. Utóbbi annyiban különbözik más növények genomi Phot szekvenciájától, hogy a fehérjekódoló részt nem szakítják meg intronok. Ráadásul a becősmohák genomjában van egy másik Phot szekvencia (ami nem egy neokróm része), ami szintén intron-mentes, így minden valószínűség miatt egy retrogén (hogy utóbbiak mik is, arról korábban már írtunk).
Mivel a becősmoha Phot szekvencia igen hasonló a becősmoha Neo génjében levő fototropin szekvenciához, az tűnik a legvalószínűbbnek, hogy ebben a fajban következett be valamiképp a Phy és Phot szekvenciák fúziója, létrehozva a Neo gént. Ami természetesen adaptív volt a mohák számára, amelyek az aljzaton élnek, így eleve kevesebb és jellegzetesen hosszabb hullámhosszú fény jut el hozzájuk. Vagyis jobb ehhez igazítani a fototropizmust is, márpedig a Phy – Phot fúzió pont erre alkalmas fehérjét hozott létre.
A páfrányok számára sokáig nem volt szüksége Neo fehérjére, ők voltak a növényvilág urai, a legmagasabbra növő szárazföldi növények. Mindez gyökeresen megváltozott a nyitva-, majd zárvatermők megjelenésével, amelyek magasabbra növő fatestet tudtak létrehozni, a páfrányokat fokozatosan visszaszorítva az aljnövényzetbe. Ebben az új élethelyzetben már adaptívvá vált a Neo jelenléte (hiszen egyre kevesebb rövid hullámhosszú kék fény jutott le hozzájuk), és valóban kb. ez az időszak az, amikor a Neo megjelent a páfránygenomokban is.
Egy másik érdekes felfedezés abból adódott, amikor különböző páfrányfajok Neo génjeit elkezdték egymással összveteni. Az így kialakuló filogenetikai fa ugyanis nagyon sok helyen másképp nézett ki, mint a rendszertani fa. Aminek az egyik oka az is lehet, hogy a Neo nem a ma élő páfrányok közös ősében jelent meg és onnan örökldődött tovább, hanem csak később került bele a páfrányok népes családjának egyik tagjába, majd onnan kellett elterjednie – hogy másképp, mint horizontális géntranszferrel (hgt-vel).
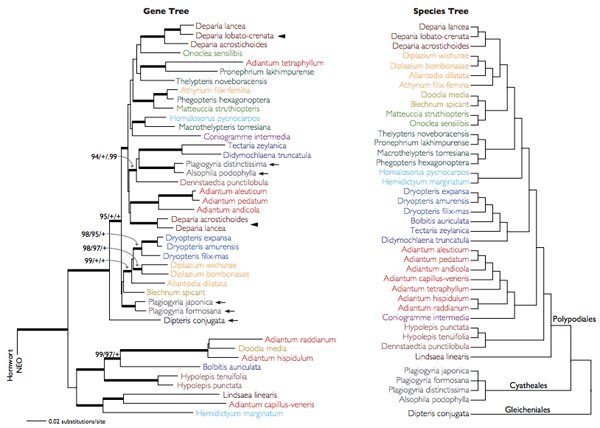
A páfrány Neo hgt-alapú születése, illetve csoporton belül való elterjedése arra utal, hogy a hgt itt nem is egy annyira ritka esemény, aminek legvalószínűbb oka a páfrányok sajátos szaporodásában rejlik. Míg a legtöbb szárazföldi növény esetében az ivarsejteket jellegzetes védőképletek veszik körbe, a páfrányok esetében az ivarsejtek nincsenek közvetlenül elzárva a külvilágtól, így könnyebben juthatnak a közelükbe más fajokból származó DNS darabok is.
Mindenestre, ez az eredmény is alátámasztja, hogy a növényi genom rendkívül dinamikus és semmiképp sem az a statikus valami, mint amilyennek ma sokan szeretnék láttatni.
Li FW, Villarreal JC, Kelly S, Rothfels CJ, Melkonian M, et al. (2014) Horizontal transfer of an adaptive chimeric photoreceptor from bryophytes to ferns. PNAS 111(18):6672-7.